Il modello di controllo
individuato nei batteri dai due studiosi francesi Jacob e Monod è conforme a
quello del controllo a retroazione positiva e riguarda in particolare la sintesi di enzimi
digestivi.
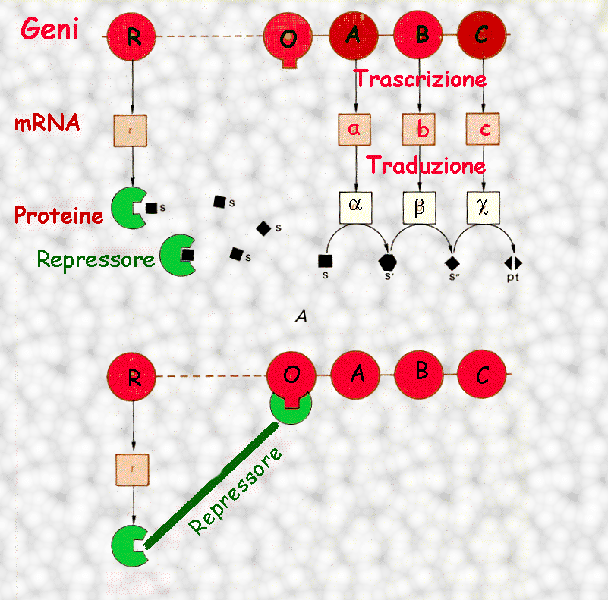
Ciascuna batteria di geni che
specifica gli enzimi che cooperano a un processo metabolico è
preceduto lungo il cromosoma da una sequenza di nucleotidi,
denominata gene operatore, che viene riconosciuta da
apposite molecole proteiche indicate col nome di repressori
(si osservino gli schemi a lato).
Se manca il substrato, gli enzimi idonei a digerirlo sono
superflui, in rapporto a ciò la trascrizione dei geni
corrispondenti viene bloccata dalla presenza di una molecola di
repressore proteico che copre il gene operatore.
Al momento in cui il
substrato penetra nella cellula, una molecola di questa sostanza
interagisce con il repressore modificandone la forma: questo si
stacca dal DNA e lascia libero il gene operatore, di conseguenza
inizia la trascrizione e la produzione dell'enzima.
L'enzima digerisce le molecole
del substrato, compresa quella che inibiva il repressore. Non
appena viene liberato, il repressore ritorna sul sito dal quale
si era staccato e inibisce la sintesi di altre molecole di
enzima, divenute ormai inutili dato che il substrato manca.
In termini di cibernetica un
processo di autoregolazione di questo tipo viene definito a
retroazione positiva con fuga a zero. Ciò che va a zero è la
concentrazione del substrato da digerire .
Questo sistema di controllo
presenta tuttavia un rischio: se l'enzima subisce una mutazione
che lo rende incapace di digerire il substrato attraverso il
quale esso viene controllato, il sistema impazzisce: basta
infatti che qualche molecola di substrato entri nella cellula
perché il gene strutturale venga attivato e non smetta più di
produrre un enzima che non serve a niente.
In effetti, nei casi meglio
studiati, si è constatato che la cellula è al riparo da questo
rischio poiché col repressore interagiscono non già le molecole
del substrato, bensì i primi prodotti della demolizione di
queste molecole .
Le cose si svolgono perciò
come se in uno stabilimento industriale la costruzione e
l'istallazione di certi macchinari e l'introduzione di certe
materie prime venisse decisa sol quando un prototipo risulta
efficiente, in caso contrario quel settore di attività viene
abbandonato.
Il controllo a retroazione negativa
sulla trascrizione.
Il dispositivo di
autoregolazione sopra descritto funziona bene per quegli enzimi
che servono solo se c'è qualcosa da digerire, altrimenti di essi
si può fare a meno. Peraltro la cellula deve
controllare in modo opportuno anche la produzione di enzimi che
sintetizzano molecole necessarie ad ogni momento, e in quantità
proporzionata a un fabbisogno variabile nel tempo. In tal caso
l'autocontrollo funziona in modo diverso:
a retroazione
negativa .
Un caso tipico riguarda la
regolazione della sintesi di amminoacidi che vengono utilizzati
di continuo, anche se in misura fluttuante, allo scopo di
sintetizzare le proteine, e che pertanto devono essere
disponibili a ogni momento nelle concentrazioni opportune.
Per rispondere a simili esigenze
la produzione dell'enzima è così governata: la macromolecola del
repressore destinato a chiudere il flusso della sintesi è
incompleta (aporepressore) e diviene funzionante solo a
patto di legarsi al prodotto finito che funge da corepressore:
nel caso considerato con una molecola dell'amminoacido sotto
controllo (fig. ).
Ad esempio: quando le molecole
dell'amminoacido Triptofano sono state sintetizzate in quantità
tali da raggiungere la concentrazione ottimale, allora alcune di
esse cominciano a legarsi ad altrettante molecole di
aporepressore trasformandole in olorepressore; questo è
capace di interagire col gene e di bloccare la sintesi del
Triptofano.
E' importante
notare che la sintesi degli amminoacidi e di altre micromolecole
complesse non avviene a opera di un singolo enzima, bensì a
opera di una batteria di enzimi: nei Procarioti i vari geni
strutturali che specificano l'intera batteria sono sempre
disposti lungo il genoma batterico uno di sèguito all'altro e
l'intera fila è controllata da un singolo gene operatore (nel
caso del triptofano gli enzimi sono cinque). E' anche da notare
che l'aporepressore acquista la capacità di interagire con
questo gene quando si è legato al prodotto finito, e non a un
prodotto intermedio della lavorazione: ciò garantisce che non si
verifichino disarmonie nel delicato processo.
La logica dell'autocontrollo nella genetica molecolare
La logica di un processo di
autoregolazione a retroazione negativa è la stessa che viene
impiegata nella gestione dei magazzini di grandi industrie:
viene misurato di continuo lo spazio disponibile per ciascun
articolo, e, non appena questo spazio è prossimo alla
saturazione parte l'ordine di arrestare la produzione o
l'acquisto di quell'articolo. Così si evitano intasamenti.
Nel caso portato ad esempio,
però, entro la cellula non è stata interrotta la fabbricazione
del prodotto finito (l'amminoacido) bensì quella del macchinario
molecolare. Ciò non costituisce un inconveniente per due ordini
di motivi. Primo, esistono dispositivi sussidiari che inattivano
gli enzimi allorché il prodotto finito ha raggiunto la
concentrazione ideale (essi non hanno rapporto diretto col DNA e
in questa sede non ci riguardano). Secondo, la riproduzione
batterica è tanto rapida che le molecole enzimatiche in eccesso
ed inattive si diluiscono (non essendo più sintetizzate) nelle
cellule della discendenza.
Sul modo in cui viene
controllata l'erogazione dell'informazione genetica nelle
cellule eucariotiche si sa poco, ma l'impegno dei ricercatori in
questo campo promette risultati interessanti a breve scadenza.
Un dato è comunque sicuro:
nella cellula eucariotica il controllo non viene effettuato su
batterie di geni strutturali disposti in fila, ma su singoli
geni. Ciò è vantaggioso poiché consente un più vario
assortimento delle molecole enzimatiche in funzione delle
necessità contingenti. Il controllo su singoli geni strutturali
implica però che una più importante quota del genoma venga
impegnata nei processi di regolazione. Infatti, mentre il
controllo diretto sui singoli geni richiede l'aumento del numero
dei geni regolatori, altri dispositivi dovranno assicurare il
loro coordinamento e la loro sincronizzazione.
Questo controllo presso gli
organismi pluricellulari si complica ulteriormente poiché
l'erogazione dell'informazione genetica viene modulata in modo
particolare per ciascun ceppo cellulare che si differenzia sia
nel corso dell'embriogenesi animale sia nel
differenziamento dei tessuti proliferativi delle piante.
Insistendo sulla similitudine
tra linguaggio umano e flusso informativo genetico, si può dire
che per la genetica batterica si conoscono bene parole e
grammatica e quindi si comprendono le frasi che definiscono il
montaggio delle molecole informazionali; anche sintassi e logica
sono conosciute, benché in modo incompleto, per cui vengono
compresi piuttosto bene l'autocontrollo e la strategia. Per
quanto concerne la genetica molecolare degli Eucarioti, invece, si deve dire che parole,
grammatica e brevi messaggi vengono compresi, mentre la
sintassi, cioè il modo con cui sono interconnessi i singoli
programmi per ora sfugge quasi del tutto.