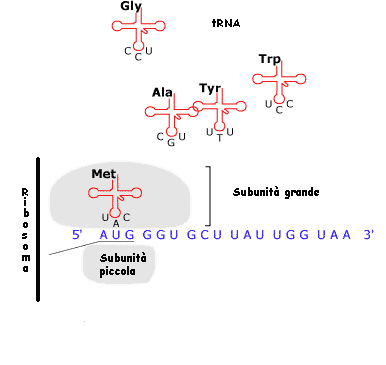
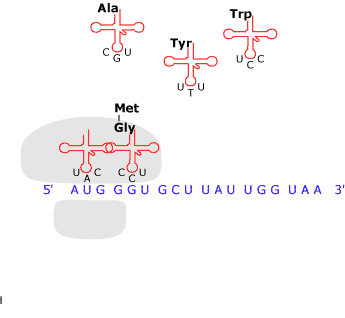
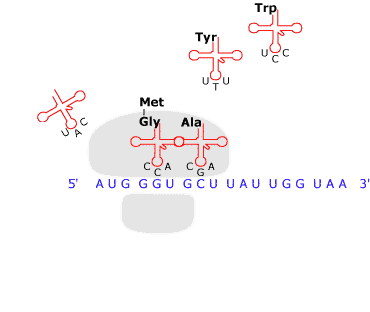
Il filamento
dell'mRNA entra in rapporto con il
ribosoma, il cui apparato, assistito
da vari tipi di molecole, procede alla lettura della sequenza
delle basi e la traduce in una sequenza di amminoacidi in
conformità con un codice, o matrice di trasformazione.
Le tappe
principali del processo sono le seguenti: presso ciascuna
tripletta di ribonucleotidi del mRNA viene trasportato
l'amminoacido 'attivato' che ad essa corrisponde in base al
codice genetico; il vettore è costituito da una speciale
molecola di RNA detta di trasporto (sigla tRNA). Dopo di ciò
viene catalizzata la formazione dei legami peptidici tra
amminoacidi adiacenti, l'energia occorrente viene fornita dalla
defosforilazione dell'amminoacido. Insomma, da una parte avanza
il filamento dell'mRNA e dall'altra esce la catena polipeptidica.
La traduzione viene effettuata
in modo rapidissimo ma, di regola solo per un numero determinato
di copie, dopodiché il messaggero viene distrutto. Questa
demolizione può sembrare uno spreco, risponde invece a un
principio generale in biologia: i messaggeri devono essere
eliminati non appena hanno assolto alla loro funzione, se ciò
non avvenisse, entro l'organismo si accumulerebbe senza freno
una congerie di molecole eccedenti al fabbisogno. L'esistenza di
un dispositivo di arresto automatico del programma non stupirà
certamente chi ha avuto occasione di manovrare una macchina
fotocopiatrice.
A causa della natura del codice,
nel quale a ciascun amminoacido corrisponde più di una tripletta
di basi del DNA diventa impossibile risalire dalla sequenza
degli amminoacidi alla sequenza di nucleotidi dell'mRNA o del
gene strutturale.
Non tutte le proteine
vengono prodotte dall'apparato sintetico con la loro struttura
definitiva. Alcune di esse subiscono modifiche che vengono
designate 'post—traduzionali'.
Tali modifiche possono
consistere nell'aggiunta di un gruppo prostetico che
conferisce alle proteine la reattività o le attitudini
fisiologiche che sono loro proprie. Nel caso dei gruppi
sanguigni ABO viene aggiunta una breve catena polisaccaridica;
nel caso delle emoglobine viene aggiunto un anello
tetrapirrolico, l'eme, che lega un atomo di Fe; nel
caso dell'emocianina, pigmento trasportatore di O2,
un atomo di Cu viene catturato dal polipeptide, e via dicendo.
Dev'essere chiaro che nella
produzione di proteine così confezionate, dopo il gene che
specifica la catena polipeptidica devono intervenire i geni che
specificano gli enzimi che sintetizzano il gruppo prostetico e
quelli che lo aggiungono alla proteina.
Altri polipeptidi diventano
fisiologicamente attivi dopo che da essi è stato amputato un
segmento: è il caso dell'insulina, della tripsina e di altre
proteine che con la loro attività danneggerebbero la cellula che
le produce: i segmenti amputati svolgono la medesima funzione della sicura
che garantisce l'incolumità a chi produce o trasporta certe
armi.
Il controllo sulla traduzione.
La regolazione dell'attività di
sintesi chimica all'interno di una cellula può essere meglio
confrontata con l'insieme dei dispositivi di controllo che
operano in una grande industria farmaceutica: la cellula è
infatti in grado di sintetizzare in dosi precise migliaia di
prodotti diversi a seconda delle esigenze funzionali del
momento.
Un programma di produzione,
comunque formulato, non risulta soddisfacente se il centro
decisionale - che nel nostro caso è il genoma stesso - non
riceve informazioni sul 'fabbisogno del mercato' che non è altro
che il fabbisogno della cellula.
Le prime conoscenze sicure sul
modo in cui il DNA riceve informazione e interviene di
conseguenza sulla sintesi proteica sono scaturite dalle ricerche
pubblicate da F. Jacob e J. Monod nel 1961, quindi poco prima
del chiarimento completo del codice genetico e delle modalità
del montaggio delle proteine. Negli anni successivi altri
studiosi hanno descritto molti altri dispositivi di
autocontrollo sicché oggi si dispone di un quadro molto ricco ed
istruttivo che però in questa sede verrà trattato in modo
succinto facendo riferimento alla genetica microbica.