Lo studio dello sviluppo embrionale è incominciato in tempi molto antichi, già in epoca classica, ma è solo nell'Ottocento che i naturalisti hanno portato l'embriologia descrittiva ad un alto livello di perfezione. In sèguito questo campo di studi è stato trascurato
poiché nel Novecento gli zoologi hanno preferito rivolgere l'attenzione all'embriologia sperimentale, argomento che non verrà trattato in questo libro. In
questa sede saranno considerate solo le nozioni generali di carattere descrittivo necessarie a comprendere varie questioni della zoologia.
La
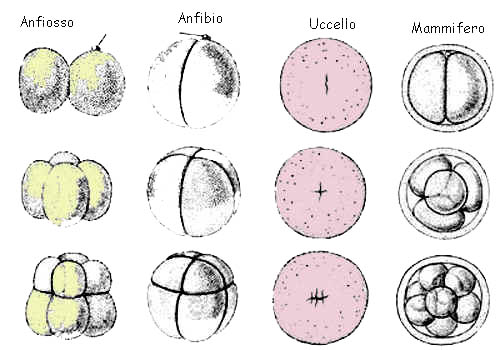
segmentazione dell'uovo che non sia sovraccarico di materiali di riserva (tuorlo) procede secondo due schemi alternativi.
Secondo uno di questi schemi i piani di divisione delle cellule che derivano dall'uovo (blastomeri) sono tra loro ortogonali (fig.) e si parla allora di segmentazione radiale. Secondo l'altro schema (forse più primitivo), i piani di segmentazione sono disposti obliquamente rispetto all'asse principale dell'uovo in modo tale che i blastomeri si sovrappongono in parte
(fig.): si parla allora di
segmentazione spirale.
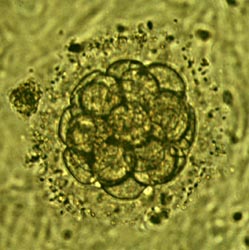
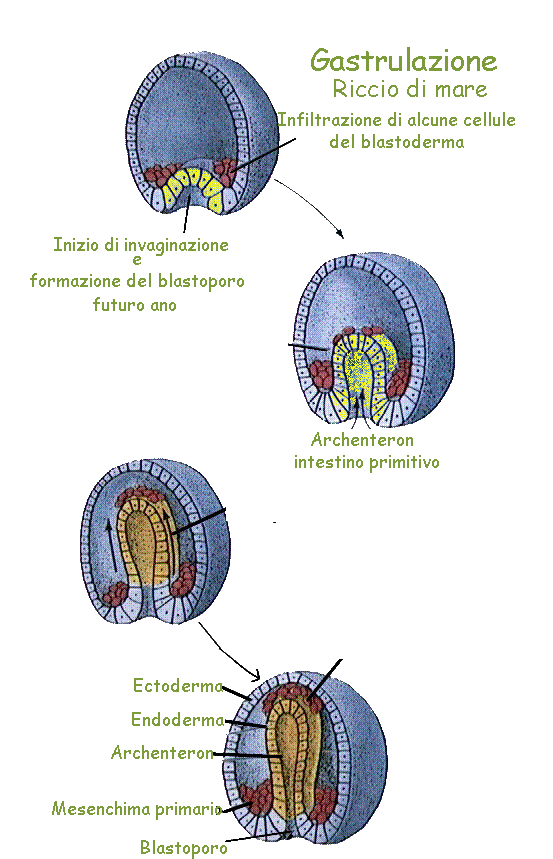
In un caso e nell'altro dopo un certo numero di divisioni cellulari l'embrione si presenta come un aggregato di cellule tondeggianti che presentano l'aspetto di una mora, gli si dà pertanto il nome di morula (fig.).
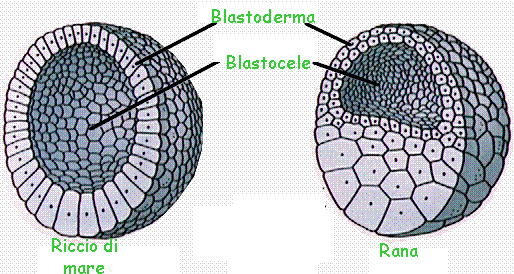
In un momento successivo all'interno della massa di cellule che costituisce la morula si forma una cavità, il
blastocele e all'embrione si dà il nome di
blastula (fig.).
Sopravviene quindi un processo particolare durante il quale la calotta di cellule che contiene maggior copia di sostanze di riserva, si introflette entro la cavità della blastula che di conseguenza tende a sparire mentre le cellule che formano la porzione superiore dell'embrione tendono a scorrere su quelle che si sono
invaginate. L'embrione a questo punto acquista la forma di un sacco a doppia parete al quale viene dato il
nome di gastrula (fig.).
Il nome di gastrula sta ad indicare che in esso è comparsa una cavità, l'intestino primitivo, che si apre all'esterno mediante un
blastoporo.
Allo strato cellulare superficiale della gastrula gli antichi embriologi hanno dato il nome di
foglietto
esterno, mentre lo strato sottostante è stato chiamato
foglietto interno. In tempi recenti il primo nome è stato tradotto in quello più oscuro di
ectoderma ed il secondo in endoderma.
Per gli animali meno evoluti lo sviluppo embrionale termina in questa fase dopo la quale si sviluppano eventuali appendici e altre strutture dell'adulto. Poiché questi organismi derivano da un embrione a due foglietti si parla di animali diploblastici (la parola è difficile, ma il concetto è semplice).
Ulteriori fasi dello sviluppo embrionale
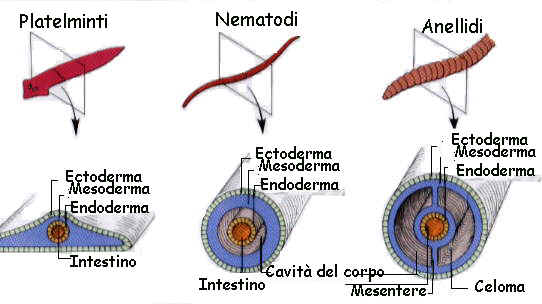
Nella maggior parte dei phyla animali il programma dello sviluppo embrionale prosegue con la formazione di un terzo strato cellulare che si inserisce tra il foglietto interno e quello esterno: ad esso viene dato il nome di foglietto medio o
mesoderma (fig.).
Il foglietto medio (mesoderma) in molti animali si sfalda presto in due strati.
Lo strato esterno tappezza la parete del corpo, quello interno fascia l'intestino; entrambi cooperano alla formazione dei mesenteri che sostengono l'intestino (fig.) all'interno nella nuova cavità. Questa cavità, detta celoma, è contenuta tra i due foglietti mesodermici.
Nella maggior parte dei phyla animali lo sviluppo embrionale termina con la formazione del terzo foglietto, e ad essi si dà il nome di triploblastici.
Negli animali triploblastici, Platelminti eccettuati, l'intestino non è più sacciforme, ma tubolare essendo munito di due aperture: la bocca e l'ano. La formazione di queste due aperture che consente un flusso di cibo molto efficiente è stato un passo evolutivo assai importante.
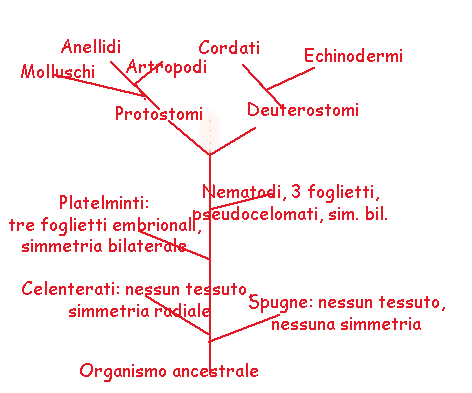
In una serie di phyla animali la bocca deriva dal blastoporo e l'ano si forma ex novo all'estremità opposta dell'embrione.
In altri phyla invece è l'apertura anale che deriva dal blastoporo, mentre la bocca si forma ex novo (fig.). I phyla della prima serie (Tab) vengono indicati col nome collettivo di
protostomi (Nemertini, Anellidi, Molluschi, Artropodi), i phyla della seconda serie sono indicati col nome di
deuterostomi
(Echinodermi, Cordati).
Istogenesi ed organogenesi
Terminato lo sviluppo embrionale il programma ontogenetico prosegue con la formazione dei tessuti (istogenesi) e degli organi (organogenesi).
L'organogenesi varia, ovviamente, in modo cospicuo a seconda delle classi e della posizione della specie nella scala evolutiva. Tuttavia si possono fornire utili indicazioni in merito all'origine di tessuti e organi, origine che rimane costante o
quasi presso tutti gli animali.
Dal foglietto esterno derivano l'epidermide e il tessuto nervoso, derivano anche i recettori di senso che mettono in comunicazione l'organismo col mondo esterno. Esoscheletro e conchiglia - presenti solo in alcuni phyla - sono strutture inerti di
produzione ectodermica.
Dal foglietto interno deriva l'intestino con le
ghiandole digestive; da esso derivano anche, in alcune classi, le strutture adibite agli scambi gassosi: i
polmoni dei vertebrati, ad esempio.
Dal foglietto medio (mesoderma) prendono origine la muscolatura, lo scheletro interno (se c'è), l'apparato escretore, nonché le gonadi. Negli animali forniti di
cavità celomatica, le lamine peritoneali sono di derivazione mesodermica.
Alla formazione di complessi organi di senso, degli arti e di eventuali altre appendici cooperano tessuti di diversa provenienza: la loro forma definitiva è conseguita attraverso la crescita differenziale ed orientata di questi tessuti.