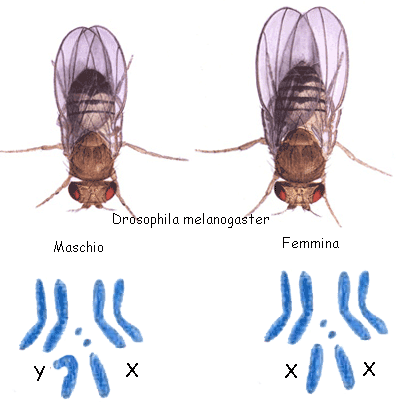
All'inizio dello scorso secolo lo studio del
corredo cromosomico rivelò che i maschi e le femmine di molte specie
di animali possono avere una coppia di
cromosomi diversi. Nei Mammiferi e in
molti insetti (nei Ditteri, ad esempio) il corredo cromosomico diploide
delle femmine risulta costituito da una doppia serie di cromosomi eguali a
due a due e nei maschi dalla stessa serie in cui però una coppia è formata
da membri diseguali.
Ai cromosomi che si presentano sempre in
coppie eguali è stato attribuito il nome di autosomi (simbolo
collettivo AA); alla coppia di cromosomi che nel maschio sono diversi è
stato attribuito il nome di eterocromosomi, assegnando il simbolo X
a quelli che sono eguali nella femmina e il simbolo Y a quello portato in
esclusiva dal maschio (la femmina ha corredo AAXX, il maschio AAXY) (fig).
Importante notare che la coppia di
cromosomi XY nel corso della meiosi non forma bivalenti e ciascun
cromosoma nel corso della anafase meiotica I passa indiviso in una delle
cellule figlie.
Negli animali così equipaggiati - tra i quali figura la specie umana - la determinazione del sesso
ha luogo secondo gli schemi mendeliani. La femmina isogametica
produce sempre e solo gameti col corredo AX mentre il maschio eterogametico produce in egual numero gameti in cui segrega il
cromosoma Y (corredo AY) e gameti in cui segrega il cromosoma X (corredo
AX). Per tal motivo le uova fecondate dall'uno o dall'altro tipo di gameti
sono in numero eguale e in numero eguale sono i maschi e le femmine .
Presso gli uccelli e
parte dei rettili, nonché presso i lepidotteri, sono eterogametiche le
femmine e omogametici i maschi, si inverte cioè la situazione.
Si modifica anche la simbologia in quanto la coppia di eterocromosomi
viene indicata con le lettere W e Z. In questi animali la formula
cromosomica del maschio è AAZZ e la formula cromosomica della femmina è
AAWZ.
Le coppie di cromosomi sessuali di cui si è
parlato nel paragrafo precedente non si equivalgono, per il fatto che solo
pochissimi geni presenti nel cromosoma X trovano il corrispondente nel
cromosoma Y e che solo una minima parte dei geni presenti sul cromosoma Z
trovano il corrispondente sul cromosoma W.
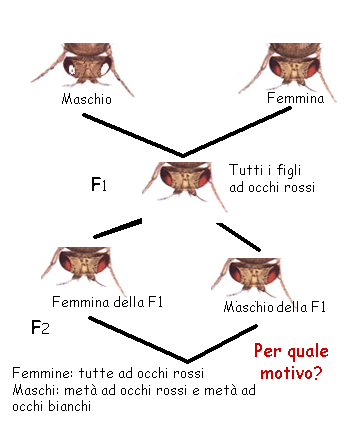
Le differenze nell'equipaggiamento
cromosomico (fig.) hanno ripercussioni importanti sulla genetica e
sulla biologia dei due sessi.
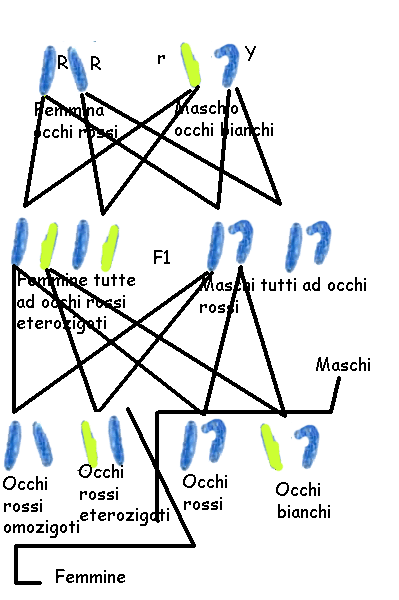
Poichè la maggior parte degli
alleli
dannosi portati in singola dose (in emizigosi) da un sesso non
risultano bilanciati da altrettanti geni non mutati presenti sul cromosoma
omologo, il sesso eterogametico risulta geneticamente più vulnerabile (regola di Haldane). E' questo uno dei motivi per cui nella specie
umana i maschi hanno vita più breve di circa 6 anni delle femmine e
possono manifestare malattie o anomalie ereditarie sconosciute nell'altro
sesso: emofilia A e B, distrofia muscolare di Duchenne, daltonismo.
Faremo in proposito il solo esempio
dell'emofilia di tipo A, provocata dall'assenza del fattore VIII per la
coagulazione del sangue.
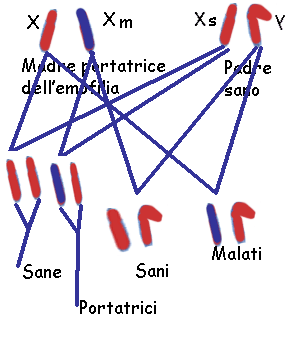
Le femmine possono essere 'portatrici sane' dell'allele per l'emofilia, che si manifesterà in metà dei loro
figli maschi, mentre una metà delle figlie femmine riceverà dalla madre l'allele
che causa questa malattia.
Fino a qualche decennio fa i maschi
emofilici morivano quasi sempre prima di raggiungere l'età della
riproduzione, sicché era praticamente impossibile che un maschio emizigote procreasse insieme a una femmina portatrice, sicché la
comparsa di femmine omozigoti per l'allele dell'emofilia è puramente
teorica. Oggi le prospettive di vita degli individui emofiliaci sono molto
aumentate grazie alla terapia trasfusionale e molti di essi raggiungono
l'età della riproduzione, ma per motivi di profilassi genetica, rinunciano
ad avere figli. Se non facessero così, e sposassero una donna non
portatrice, tutti i loro figli maschi nascerebbero indenni poiché il solo
cromosoma X che essi possiedono proviene sempre dalla madre, mentre tutte
le figlie femmine sarebbero portatrici sane dell'allele per l'emofilia
(fig.); esse infatti ricevono necessariamente uno dei due cromosomi X
del padre.