Scelta e regolazione delle
traiettorie
Non
è errato affermare, parafrasando un antico proverbio, che i
bisogni sono il motore di ogni comportamento. I bisogni sono
molto diversi a seconda delle esigenze proprie della specie e
delle situazioni particolari in cui si trova l'individuo, c'è
però un fatto che li accomuna tutti: la necessità di
raggiungere la fonte ove può essere ristabilito
l'autocontrollo di un parametro biologico, la necessità cioè di
raggiungere il luogo ove il bisogno può essere appagato,
bevendo, riscaldandosi, nutrendosi, e via dicendo.
A tal fine l'organismo deve
individuare il segnale proveniente dalla fonte e seguire una
adeguata traiettoria per raggiungerla.
Se dalla fonte non parte
alcun segnale che l'animale possa utilizzare, l'unica
strategia possibile è quella di muoversi a caso. Ad esempio
quando si solleva e rovescia una grande pietra si espongono al
calore e alla luce del sole decine di animali che stavano
rifugiati là sotto, e che rischiano di morire disidratati o
vittime di predatori di passaggio. In queste circostanze i
Vertebrati ben equipaggiati con organi di senso e
memoria si
dirigeranno verso un altro rifugio a loro noto, ma la folla di
artropodi si muoverà in direzioni scelte a caso, rallentando
però la fuga a mano a mano che la luce si attenua e l'umidità
cresce (chinokinesi): i
più fortunati, e resistenti, si ritroveranno sotto la protezione
di un altro sasso, di un tronco, in una buca.
Recezione
ed elaborazione di segnali chimici
Le molecole di una sostanza
solubile che si trova sospesa nel solvente, nel passare in
soluzione, si espandono, a partire dall'origine, secondo i raggi
di una sfera (o di una emisfera nel caso che la sostanza che si
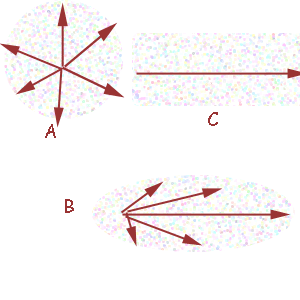
scioglie riposi sul fondo). Il fuoco di massima concentrazione
sarà situato al centro della figura geometrica (fig. 2, A). Questa
proprietà, ovviamente, riguarda anche le sostanze che fungono da
segnale per un qualche organismo. Sicché un vivente che tenta di
raggiungere la fonte di quello stimolo deve necessariamente
risalire il gradiente di concentrazione.
Se però nell'ambiente vi è un
flusso direzionale, allora le molecole si distribuiranno secondo
un ellissoide, o un mezzo ellissoide, allungato e la massima
concentrazione (e fonte dello stimolo) si troverà nel fuoco
dell'ellissoide situato a monte. Al limite, se il flusso è
rapido (in un fiume, in una località battuta da vento costante)
le molecole si distribuiscono secondo una sorta di tunnel, ad
una estremità del quale si trova la fonte (fig. 2, B, C).
I modelli
cibernetici della
omeorresi permettono di chiarire che nei primi due
casi la traiettoria che un organismo segue per raggiungere la
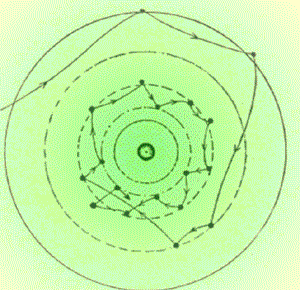
massima concentrazione risponde a una regola molto semplice: non
appena l'intensità del segnale si riduce, sterzare di un angolo
fisso (fig. 3). Se l'ambiente non è turbolento, la traiettoria
apparirà come una linea spezzata simile a una grossolana spirale
Il programma da
effettuare appare molto semplice, tuttavia la faccenda presenta
un punto critico: il sensore segnala il numero di molecole con
cui esso interagisce nell'unità di tempo, mentre la sterzata
dev'essere comandata da una variazione in meno di questo numero
(cioè dalla derivata negativa dell'intensità del segnale). Ciò è
possibile solo se viene effettuato un confronto tra il segnale
all'istante e il segnale ricevuto un momento prima. Che questo
confronto possa avere luogo all'interno di una cellula è del
tutto verosimile, cosa avvenga in realtà è per ora solo oggetto
di ipotesi.
La recezione della luce e
l'orientamento conseguente
La
luce è tra le forme di energia più diffuse in questo nostro
mondo, e da essa dipendono, ovviamente, tutti gli organismi
fotoautotrofi; da essa dipendono però in un qualche modo anche
gli altri organismi i quali o la ricercano, o la fuggono.
Presupposto per la percezione della luce è che esista un
fotopigmento che si decompone alla luce. Tale decomposizione
libera energia, e questa energia può innescare un qualche
comportamento. Un fotopigmento presente dai batteri fino ai
Vertebrati è la rodopsina. La molecola di questa sostanza è
formata da un'aldeide (derivata da un idrocarburo carotenoide)
di color rosso, il retinale, e da una proteina, la
opsina, la cui struttura si conserva, sia pure con molte
variazioni, da un capo all'altro della scala dei viventi. Sotto
l'azione della luce il retinale cambia forma distaccandosi in
parte, o del tutto, dal supporto proteico e ciò genera un
segnale interno, di natura elettrica; appositi enzimi ridanno
alle molecole di rodopsina la loro forma primitiva a mano a mano
che esse si distaccano. Se però il processo di decomposizione
prevale su quello di resintesi si verifica l'abbagliamento
del recettore (o dell'occhio); l'abbagliamento è analogo alla
saturazione che rende inutilizzabili i recettori olfattivi
in un ambiente saturo di una sostanza odorosa.
Gli organismi unicellulari, come
l'euglena della figura, dispongono di un sensore impari
schermato in modo da ricevere la luce solo da una parte; gli
organismi pluricellulari hanno organi visivi pari, schermati
allo stesso modo.
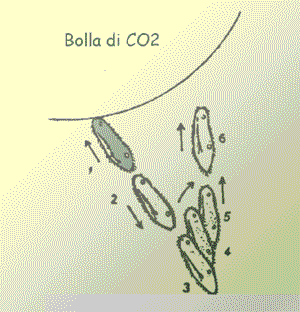
Il dirigersi verso la luce (fototassi
positiva) è guidato, in tutti i microorganismi muniti di un
solo sensore, da un programma molto semplice: "se l'intensità del
segnale diminuisce, sterza di un angolo fisso". Il fuggire della
luce (fototassi negativa) è sorretto da un programma
inverso: "se l'intensità del segnale aumenta, sterza di un angolo
fisso".
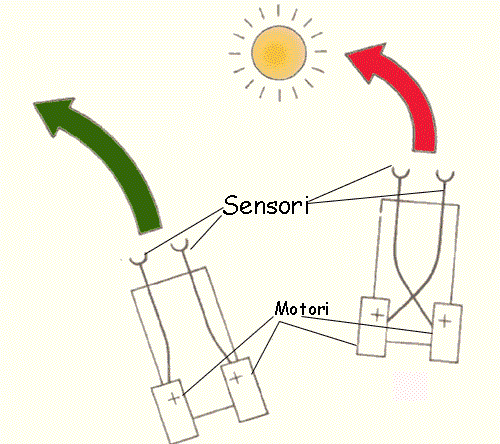
Presso gli
organismi muniti di due fotorecettori o di due occhi pari il
programma per dirigersi verso la luce, dipende dal confronto tra
l'intensità registrata da una parte e dall'altra: "se vi è
discrepanza l'organismo sterzerà dalla parte del recettore che
riceve il segnale più forte".