Strategie vincenti
A
ogni predatore per raggiungere la vittima occorre in primo luogo
reperire le tracce, i segnali, che questa - suo malgrado -
lascia dietro a sè. L'animale che non dà alcuna informazione di
sè dovrebbe essere al sicuro.
Tuttavia anche animali
apparentemente elusivi, come i calamari giganti che vivono nei
bui abissi marini e le farfalle che volano silenziose
nell'oscurità notturna hanno "nemici" pericolosi dai quali non
sono adeguatamene protetti. Capodogli, pipistrelli e altri
predatori riescono a raggiungere la preda meglio nascosta -
sondando l'ambiente mediante l'emissione di qualche forma di
energia da loro generata di cui raccolgono il riflesso, o l'eco.
Proprio come fa un ladro col fascio di luce di una torcia
elettrica, come fa un medico che sottopone il paziente
all'ecografia, come fa un cieco che procede battendo il bastone
sul selciato.
Ma forse conviene ricorrere a
esempi tratti da competizioni più cruente e drammatiche.
Durante la seconda guerra
mondiale erano in uso due diversi sistemi per individuare nel
buio della notte gli aereoplani nemici.
Presso un esercito si
impiegavano due enormi imbuti mobili in senso verticale e
orizzontale che raccoglievano e amplificavano il rumore dei
motori proveniente dal cielo: di conserva si muovevano le
fotoelettriche (spente) e le artiglierie. Al momento in cui il
segnale captato da entrambi gli imbuti coincideva e aveva
raggiunto il valore massimo, si accendevano le fotoelettriche e
i cannoni si trovavano già puntati sul bersaglio illuminato.
L'altro esercito impiegava
un'apparecchiatura rotante che lanciava impulsi di onde-radio
ultracorte e ne raccoglieva l'eco riflesso dall'aereo che era
stato investito da questi impulsi. Distanza, traiettoria e
velocità venivano identificate su una schermo a raggi catodici,
e nel contempo le artiglierie venivano puntate in funzione di
questa identificazione.
Il primo sistema di
identificazione e puntamento presentava uno svantaggio: il
segnale acustico poteva risultare distorto dalla disposizione e
dal movimento degli strati d'aria e - soprattutto - il pilota
dell'areo nemico veniva informato dell'imminente arrivo dei
proiettili dal fascio di luce che lo lambiva; ciò gli dava la
possibilità di scampare picchiando, ad esempio, velocissimo
verso il suolo dove non poteva essere seguito.
Il sondaggio dello spazio con
fasci di energia elettromagnetica di cui si raccoglieva l'eco
era molto più efficace e consentiva alle artiglierie di sparare
al buio, attraverso le nuvole, con alta probabilità di
raggiungere l'aereo di sorpresa.
Solo dopo questi sviluppi della
tecnologia militare ci si è accorti che fin da epoche
remotissime varie sorta di animali rintracciano la preda e
scansano ostacoli sondando lo spazio con sistemi del tutto
simili.
Sondaggio mediante
ultrasuoni
I
pipistrelli (microchirotteri) lanciano onde
ultrasonore che vengono riflesse dagli ostacoli fissi o mobili.
In base all'analisi di questi echi, tali animali anche nelle
notti più buie e nelle caverne più profonde sanno evitare gli
ostacoli fissi, e sanno stabilire una rotta di collisione con
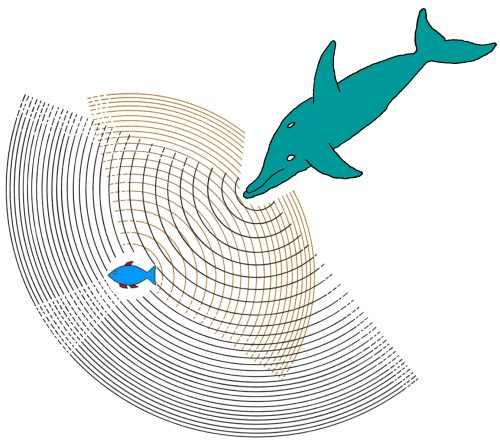
gli insetti da abboccare . Il potere risolutivo del
dispositivo di ecolocazione è tanto maggiore quanto più è
alta la frequenza dei suoni emessi; la frequenza usata dai
pipistrelli (da 20 a 90 kiloHertz) è tale da consentire la
percezione a un metro di distanza di fili di solo 0.07 mm di
spessore (inferiore a quello dei capelli) e di abboccare
moscerini e farfalle aventi sezioni di urto di 2 mm2 e anche meno (fig).
E' da notare che alcune specie
di farfalle notturne hanno recettori sintonizzati sulle
frequenze più usate dai pipistrelli nelle loro cacce (30-45
kiloHertz) e che non appena percepiscono il segnale lanciato del
predatore, si difendono con lo stratagemma illustrato sopra: si
lasciano cadere a picco. Altre farfalle hanno un
rivestimento che assorbe tali onde attenuando così il pericoloso
segnale di ritorno che le rivela al predatore. Altre ancora,
emettendo segnali sulla lunghezza d'onda usata dal pipistrello,
gli comunicano che sono cariche di veleno e che è meglio per lui
lasciarle campare.
Presso i Cetacei esistono
sistemi analoghi che consentono di evitare gli ostacoli durante
la navigazione e di localizzare le prede nella vastità del mare.
Emettono però onde ultrasonore con frequenza molto più elevata
di quelle del pipistrello (fino a 200 kiloHertz) per compensare
la più alta velocità di conduzione del suono posseduta
dall'acqua (circa quintupla di quella dell'aria); gli odontoceti
(delfini e capodogli), inoltre, sono in grado di far convergere
queste onde in fasci piuttosto ristretti mediante una lente
formata da una massa globulare di olio situata tra il cranio e
la mascella .
Simile attrezzatura consente al
capodoglio di localizzare i calamari giganti (Architeuthis),
di cui si diceva prima, che vivono a 400-600 metri di
profondità. Sembra anche, ma non se ne ha la certezza, che il
capodoglio riesca a paralizzare la vittima lanciandole contro da
vicino una salva di ultrasuoni di spaventevole intensità.
L'exploit più
straordinario, tuttavia, è quello dei delfini di fiume (Platanistidi)
i quali, vivendo in acque perennemente torbide, hanno perso
l'uso della vista (l'olfatto l'avevano perso molto prima) e
dipendono in tutto e per tutto dai segnali acustici che lanciano
sia per evitare gli ostacoli, sia per localizzare i pesci di cui
si nutrono.
Il sondaggio
dell'ambiente con campi elettrici
I pesci che si muovono nelle libere acque del mare o dei laghi,
cioè i pesci pelagici, incontrano pochi ostacoli, ma quelli che
vivono sul fondo, e si debbono muovere in acque fangose o al
buio, rischiano continue collisioni con scogli o con altri
animali pericolosi
Questo problema ha trovato una
soluzione insolita nel regno animale. Il pesce produce, mediante
un tessuto neuromuscolare specializzato, una rapida successione
di scariche elettriche che generano campi elettrici.
Se l'ambiente è elettricamente
isotropo, questi
campi risulteranno simmetrici e di forma regolare;
se però in esso compare un corpo cattivo conduttore (un sasso) o
un corpo buon conduttore (un pesce) il campo viene distorto in
modi diversi. Appositi sensori (recettori elettrici) dislocati
sui fianchi dell'animale registrano tali variazioni e l'animale
si regola di conseguenza evitando ostacoli ed eventuali
aggressori, e identificando le prede.
In acqua dolce, dotata di bassa
conducibilità (poiché gli elettroliti e quindi gli ioni
disciolti sono pochi), il sistema funziona bene anche a bassi
voltaggi e sono molti i pesci di fondo che se ne servono.
Tuttavia la scarica del tessuto neuromuscolare di alcune specie
può raggiungere un'intensità (fino a 1A) e un voltaggio (fino a
600 V) tali da poter tramortire la preda o l'aggressore. E'
questo il caso della ben nota anguilla elettrica dei fiumi
dell'America tropicale (Electrophorus electricus) e di un
pesce gatto (Malapterurus electricus) dei bacini
dell'Africa equatoriale (fig. 9-17).
Nei mari, ove
l'acqua porta in soluzione molti elettroliti e ha quindi una
conducibilità molto elevata, si formano campi elettrici deboli e
molto ampi, poco utili per dirigere la navigazione. I pesci
marini che generano campi elettrici sono selaci di fondo, le
torpedini, che in un remoto passato dimoravano nelle acque
interne e se ne servivano per la navigazione, mentre oggi se ne
servono per difesa e per offesa. Solo una specie di teleosteo
elettrico vive in mare e i suoi segnali pare servano per
comunicare con i propri simili. (Interessante ricordare che il
primo rapporto dell'uomo con l'elettricità è stato appunto con i
pesci elettrici, e che Volta e Galvani ne hanno fatto oggetto di
attento studio).
Il comportamento del cane da
caccia
Il comportamento animale si
complica ovviamente quando gli organi di senso si fanno più vari
ed efficienti.
Porteremo ad esempio il caso di
un cane affamato a caccia di lepri (si badi che di regola il
cane caccia in branco). Il cane fiuta inquieto il terreno e il vento. Il vento gli porta
l'odore di una miscela di molecole che gli è ben nota, si mette
dunque in moto procedendo contro vento. Sono i recettori termici
della pelle umida che circonda le narici che gli permettono di
identificare la provenienza del vento e quindi dell'odore. Il
cane fiuta il vento e il terreno spostandosi da destra verso
sinistra e, quando l'odore si attenua, torna a portarsi verso
destra finché il segnale si attenua ancora, e così avanti a
zig-zag finchè i recettori olfattivi sono tutti saturati da
quelle molecole: ciò indica che la preda è vicina.
L'animale ora 'punta' silenzioso
utilizzando l'udito, e, quando ode il cauto sfrascare della
lepre si lancia tra l'erba: la lepre fugge via. Il cane ora
insegue a vista. La lepre descrive un ampio semicerchio, ma
l'inseguitore non tenta alcuna scorciatoia, non taglia secondo
la corda dell'arco di cerchio poiché la preda può scartare di
lato lasciandolo spiazzato. La lepre è ora fuori di
vista al coperto di un rilievo. Il cane ricomincia il programma
da capo: prima fiutare, poi ascoltare, e quindi inseguire a
vista, sempre con la speranza di utilizzare anche il tatto e il
gusto.
Questa sequenza ha una sua
logica: individuare prima i segnali olfattivi che persistono nel
tempo e seguirli fin dove è possibile, raccogliere poi i segnali
acustici che forniscono un più pronto orientamento, pur
aggirando gli ostacoli; dopo l'udito conviene usare la vista, la
quale fornisce segnali direzionali (ma che vengono intercettati
da ostacoli), puntando sempre diritto sull'obiettivo.
Il territorialismo
è un comportamento molto diffuso, consistente nell'escludere
competitori - specialmente quelli della propria specie e del
medesimo sesso - da una determinata area di foraggiamento e
riproduzione. I confini dell'area protetta vengono di solito
marcati con sostanze odorose - secrezioni
ghiandolari ed escrementi - , ma molti uccelli essendo poveri di
olfatto inibiscono l'accesso ai concorrenti con il loro canto.
In questo caso il canto può fungere anche da invito alla
femmina.
Il territorialismo
verrebbe designato da una certa corrente di pensiero come
comportamento 'egoistico', ma l'uso di questo termine sarebbe in
questo caso oltre che improprio anche grossolanamente errato.
Il comportamento territoriale
infatti ha l'effetto di inibire la riproduzione ad un
contingente della popolazione al fine di evitare un
sovraffollamento e quindi la spoliazione del territorio con
danno grave per le generazioni future.
I carnivori manifestano una
sorta di territorialismo anche nei riguardi di carnivori di
specie diversa, le contese tra cane e gatto ne sono un esempio.
A proposito dei gatti va poi aggiunto che la loro riluttanza a
lasciare una dimora vecchia per una nuova è legata anche al
fatto che nella prima avevano difeso (e talvolta conquistato a
fatica) un proprio territorio che abbandonano molto mal
volentieri, dovendo ricominciare tutto da capo.
Molti animali conducono vita
gregaria. Le mandrie di grossi erbivori, gli sciami di pesci,
'le scuole' dei cetacei, gli stormi di pappagallini ondulati e
di stornelli ne offrono esempio.
La vita di branco
offre vantaggi soprattutto per la difesa: la tigre teme di venir
calpestata a morte dalla mandria di bufali atterriti, e il
leopardo evita di assalire un branco di babbuini che si muove
con i maschi più robusti disposti all'esterno,
mentre l'antilope 'diluisce' nel branco la probabilità d'essere
presa di mira in modo particolare.
Per tale motivo la
vita di branco è evoluta presso i più vari gruppi animali
imponendo alcune soluzioni parallele e altre divergenti. La gerarchizzazione è un elemento costante della vita di branco dei
vertebrati superiori per cui gli individui sono subordinati
l'uno all'altro e tutti lo sono all'individuo
a, il capo del branco. Questi ha il compito di tenere unito il
gruppo e di guidarlo e anche di intervenire per sedare le risse.
La gerarchia non è stabile: le malattie e le ferite decretano
spesso la retrocessione del capo, mentre i giovani, a mano a
mano che acquistano vigore ed esperienza, avanzano nella
gerarchia. Presso alcune specie l'avanzamento può essere
scandito da combattimenti che culminano con lo spodestamento
dell'individuo
a.
Il combattimento a volte è cruento, ma più spesso è ritualizzato;
i contendenti si limitano a 'mostrare i muscoli'. Nei primati la
posizione gerarchica della femmina può variare anche in rapporto
alla fase del ciclo riproduttivo.
Altra
caratteristica della vita di branco è il reciproco
riconoscimento dei membri (anche se i modi sfuggono
all'osservatore umano): due branchi molto numerosi si possono
mescolare all'abbeverata, per esempio, e quindi si separano
senza alcun errore.
Nei branchi di ungulati, di pinnipedi (foche, otarie), e presso talune specie di uccelli
l'accesso alle femmine recettive è riservato al maschio
dominante il quale si costituisce una sorta di harem, per cui
buona parte della nuova generazione è costituita dai suoi figli.
Nei branchi dei primati non
umani non avviene altrettanto: di solito le femmine in estro
manifestano all'inizio una certa promiscuità che non provoca
gelosia; in sèguito possono formarsi coppie stabili per qualche
tempo, ed i soggetti vengono indicati come 'consorti'.
La vita di branco
non modifica il rapporto tra madre e figli, che spesso ne viene
rafforzato, e allo stesso tempo promuove lo sviluppo dell'eredità
culturale: molte attività che in altre specie sono esplicate
in modo puramente istintivo nelle specie gregarie vengono
apprese osservando i propri simili, e ciò risulta vero persino
per le cure materne: una scimmia che sia stata allevata fin da
neonata senza contatti coi suoi simili manifesta anomalie di
comportamento tra le quali l'incapacità di accudire al proprio
neonato.
Talvolta gli individui di specie
gregarie presentano una qualche forma di territorialità
riservandosi uno spazio all'interno del branco, non si osserva
però una territorialità di branco.
La vita gregaria
presso alcune specie si è evoluta culminando nella vita sociale.
E' difficile stabilire il confine tra i due modi di vita, e il
caso dei gruppi di scimpanzè ne offre un esempio. In linea di
massima si può dire che un animale sociale non è in grado di
sopravvivere a lungo se rimane del tutto isolato e si può
aggiungere che in una società vi è divisione dei compiti tra i
membri. Ma nel caso dei mammiferi ciò non risulta sempre vero.
Costumi sociali sono comparsi
presso le tèrmiti (insetti isotteri) presso le vespe, le api e
le formiche (insetti imenotteri), presso i canidi e i primati.
Se ne farà cenno nei capitoli dedicati a tali animali. In questa
sede è però necessario segnalare tre fenomeni che si sono
affermati a sèguito della evoluzione della vita sociale.
In primo luogo, sono inibiti
tanto il territorialismo individuale quanto l'aggressività tra
membri del gruppo, comportamenti che disgregano la struttura
superindividuale.
In secondo luogo,
le comunità di animali sociali tendono a comportarsi ai fini
dell'evoluzione, come 'superindividui'. Sicchè tra queste
comunità si può istituire una competizione che porta alla
selezione di caratteri del tutto inconsueti o impensabili presso
gli individui di animali non sociali: la sterilità, ad esempio,
di una quota di individui. Ad esempio un nucleo
sociale che ha un tasso riproduttivo tale da deteriorare
l'ambiente in cui vive sparisce, mentre un altro nucleo, che per
motivi fisiologici o comportamentali ha un tasso riproduttivo
inferiore sopravvive ed eventualmente si moltiplica, finendo col
sostituire quello che appariva avvantaggiato dalla maggiore
prolificità.
In terzo luogo,
l'attività riproduttiva degli animali sociali tende a
modificarsi fino al punto che, presso le società di insetti, e
presso certi roditori che vivono sottoterra (gli Eterocefali),
la riproduzione è riservata a una sola coppia fertile; ciò ha
probabilmente lo scopo di evitare i danni causati dall'endogamia.
Infatti gli accoppiamenti o la costituzione della coppia
avvengono al momento della sciamatura, quando le femmine fertili
abbandonano la comunità in cui sono nate e si incontrano con
maschi provenienti da altre comunità.
Le emozioni hanno sempre
attirato l'attenzione degli psicologi e dei moralisti per il
loro aspetto incoercibile : l'ilarità fa esplodere risate
irrefrenabili e la collera può far compiere furiose aggressioni
a una persona mite, reazione del tutto contraria ai suoi
principi.
Descartes
(1596-1650) ha dibattuto la questione in termini scientifici e
duecent'anni dopo Claude Bernard (1813-1878) l'ha trattata sotto
il profilo fisiologico. Nei decenni seguenti se n'è parlato poco
poiché agli studiosi di psicologia comparata è sembrato un
arbitrio l'attribuire agli animali le emozioni cui va soggetto
l'uomo. Finita la seconda guerra mondiale il comportamento
aggressivo è stato portato al centro dell'interesse, poiché era
diventato drammaticamente chiaro che esso poteva portare alla
fine dell'intera umanità.
Per orientarsi sui rapporti tra
comportamento e fisiologia un filo conduttore può essere offerto
- una volta di più - dal confronto tra la tecnologia del vivente
e la tecnologia delle macchine.
Claude Bernard ha
saputo dimostrare, nel secolo scorso, che il susseguirsi di
reazioni fisiologiche che accompagnano l'insorgere della paura e
della collera ha lo scopo di mobilitare tutte le risorse
dell'organismo. Le reazioni vasomotorie provocate dalla scarica
di adrenalina sono numerose e coordinate : si riduce la
circolazione cutanea attraverso la vasocostrizione periferica
(pallore) mentre aumenta l'irrorazione dei muscoli e di alcuni
organi interni. La spremitura della milza accresce la massa
degli eritrociti circolanti mentre la dilatazione dei bronchi
agevola la ventilazione polmonare e quindi gli scambi gassosi,
poiché nel contempo aumenta la gittata cardiaca. Aumenta anche
la coagulabilità del sangue, circostanza che, insieme con la
vasocostrizione periferica, limita le conseguenze di eventuali
ferite superficiali. Parallelamente si riduce la sensibilità
dolorifica e cresce la rapidità delle risposte motorie.
Il dispositivo
messo in luce dal grande fisiologo francese ha trovato imitatori
(involontari) presso i costruttori di motori. Alcuni autocarri
sono muniti di un dispositivo che accresce la potenza del motore allorchè il veicolo deve affrontare una forte pendenza a pieno
carico; anche il motore (a pistoni) di certi aerei da caccia era
munito di un dispositivo di sovralimentazione da inserire in
vista di un combattimento. Superata l'emergenza il camionista o
il pilota disattivano il dispositivo per non rovinare il motore.
Anche la macchina fisiologica, finito il pericolo, ritorna al
ritmo di base: star sempre sotto pressione riduce le prospettive
di vivere a lungo.
Il nodo
dell'intera questione sta nel capire come l'organismo registra in modo non consapevole e non
ragionato l'emergenza, o il pericolo, e scatena la mobilitazione
generale, mobilitazione del tutto fuori dal controllo
volontario.
Non è errato e nemmeno
semplicistico ammettere che durante l'esecuzione di un programma
il sistema nervoso commisuri, in modo del tutto inconscio, le
risorse fisiche alle difficoltà che si susseguono: allorquando
un ostacolo difficilmente superabile si para dinnanzi, questo
sistema provoca la scarica di adrenalina e con essa il corteo di
reazioni fisiologiche sopra descritte: il soggetto prova
collera. Sperimentare in proposito non è difficile (ma non
sempre consigliabile) : basta intralciare chi sta eseguendo un
lavoro urgente e delicato, basta ostacolare un autista che sta
eseguendo un sorpasso, per vedere questi soggetti in preda alla
collera, per renderli aggressivi.
Anche l'insorgere della paura
trova spiegazioni semplici : ogni e qualunque programma è
governato dall'afflusso di certe informazioni sensoriali; se
qualcuna di esse viene meno, la paura insorge automaticamente.
Sorge ad esempio quando si sente mancare la terra sotto i piedi
mentre si corre, quando un improvviso frastuono nasconde un
fievole richiamo che si sta seguendo, quando non si rintraccia
nell'archivio mnemonico la risposta da dare a un quesito. La
reazione fisiologica alla paura è duplice. Da una parte
l'arresto del programma, e quindi l'immobilizzazione (se
possibile), dall'altra la ricerca di un percorso alternativo e
la fuga.
Naturalmente gli esempi esposti
forniscono solo un filo conduttore per la comprensione degli
stimoli che innescano le emozioni. Non occorre spingere oltre
questo esame; in questa sede basta aver chiarito che
l'insorgenza delle emozioni è al di fuori del controllo
volontario. Ciò comunque non esclude che un controllo volontario
possa inibire, non le reazioni fisiologiche, ma le reazioni
aggressive, o, quantomeno, che possa agire sulla misura delle
reazioni aggressive.