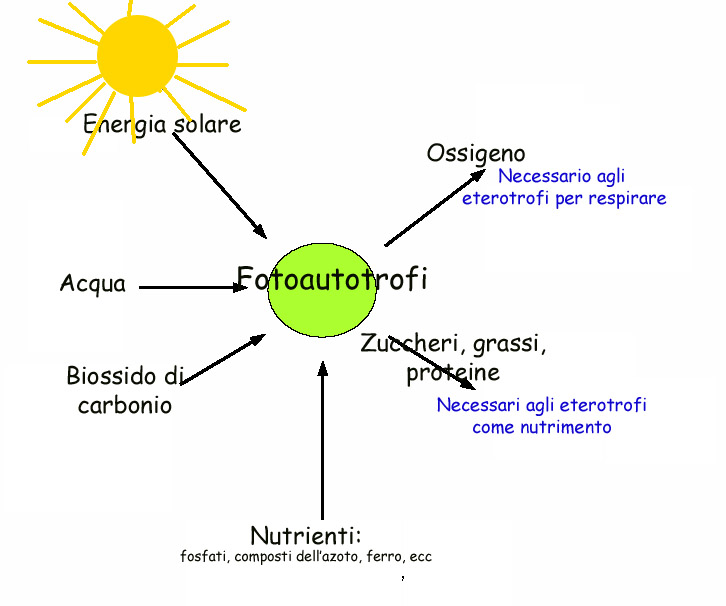
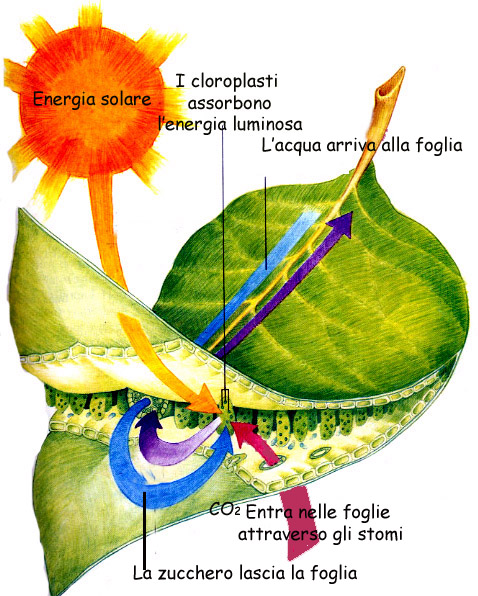
Prenderemo ora in considerazione il flusso di
energia che percorre gli organismi autotrofi grazie alla fotosintesi
clorofilliana.
La fotosintesi è un processo biochimico
alquanto complicato che è stato chiarito nelle sue grandi linee tra la fine
del Settecento e l'inizio dell'Ottocento. Il processo venne dapprima riassunto
nella reazione :
6 CO2
+ 6H2O
+ luce
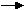 C6 H12O6
+ 6 O2 [1]
C6 H12O6
+ 6 O2 [1]
che si legge: nel corso della fotosintesi
l'anidride carbonica interagisce con l'acqua e, grazie all'apporto energetico
della luce e alla catalisi operata dalla
clorofilla si trasforma in glucosio
con liberazione di O2 .
Ricerche più recenti hanno dimostrato che la
reazione base ha andamento più complesso e che conviene scrivere la
formula riassuntiva nel modo seguente :
6 CO2
+ 12 H2O
+ luce
C6H12O6
+ 6 O2
+ 6 H2O
[2]
allo scopo di mettere in evidenza che
l'ossigeno che si libera durante la fotosintesi deriva dalla scissione di
molecole d'acqua e non è quello contenuto nelle molecole dell'anidride
carbonica.
Tuttavia neppure questa seconda formula
riassuntiva dà una soddisfacente idea dell'intero processo fotosintetico, che
è stato chiarito anche nei suoi aspetti energetici solo intorno al 1965. E'
opportuno in questa sede entrare in particolari, non per il gusto di allineare
formule chimiche, ma per chiarire il modo in cui procedono certi processi di
biochimica dinamica all'interno delle cellule.
Protagonista della fotosintesi è la
clorofilla, la cui molecola è formata dal grosso e complicato anello policiclico delle porfirine dal quale si diparte una lunga catena
idrocarburica .
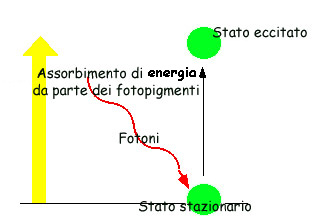
La clorofilla attivata dalla luce catalizza la
reazione :
H2O +
NADP + ADP + P + fotoni NADPH– + ATP + H+ + 1/2O2+e- [3]
Questa reazione si legge così: la luce provoca la scissione
dell'acqua nei suoi componenti nonché l'ionizzazione dell'H in H+
più elettroni e-
i quali trasformano l'accettore di elettroni NADP in NADPH-; l'energia liberata provoca inoltre la fosforilazione dell'ADP in
ATP.
Il potenziale riduttivo del NADPH- e
l'energia immagazzinata dell'ATP azionano poi la macchina della riduzione
della CO2 .
L'aspetto energetico della
fotosintesi può essere riassunto anche nel modo seguente: la clorofilla
eccitata dai fotoni si comporta come una pompa che sottrae elettroni a un
datore (l'acqua) per trasferirli a un accettore: il flusso di elettroni così
generato accumula energia e potenziale riduttivo rispettivamente nei sistemi ADP
/ATP e NADP / NADPH.
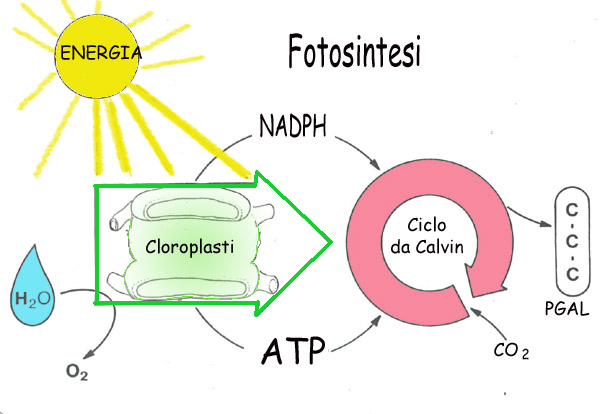
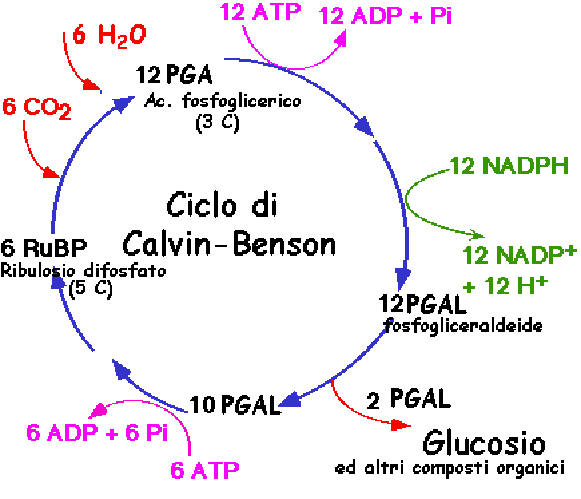
L'energia accumulata nel legami degli ATP e
dei NADPH, viene utilizzata per costruire molecole organiche in un ciclo che
prende il nome di Ciclo di Calvin-Benson, dai loro scopritori. La figura riassume l'andamento
energetico e chimico della fotosintesi ed indica i nomi e le caratteristiche
di un certo numero di sostanze chimiche che intervengono nelle varie tappe del
processo. Non è indispensabile mandare a mente i nomi e le proprietà di tutti
i protagonisti, è però importante cogliere il quadro d'insieme e rendersi conto
che l'organizzazione dei catalizzatori rende l'apparato fotosintetico molto
simile a un macchinario molecolare minutissimo ed efficientissimo: quando il
bisogno energetico della cellula aumenta essa risponde, non già aumentando le
dimensioni dei macchinari (come fanno certe industrie create dall'uomo), ma
moltiplicando i macchinari stessi, cioè le macromolecole enzimatiche e gli
organuli che le contengono.
La fotosintesi nei Batteri
procede in modo alquanto diverso da quello qui descritto, poiché in essi la
clorofilla non catalizza la scomposizione dell'acqua, bensì la scomposizione
di H , SH e di composti organici del tipo H2A.
In questi casi non viene prodotto ossigeno che ha azione nociva sui componenti
del citoplasma di questi organismi. Nei
Cianobatteri, e nei Chlorobi invece,
i quali dispongono di un dispositivo che neutralizza gli effetti nocivi
dell'ossigeno, la fotosintesi procede come sopra descritto.
Fotosintesi e composizione
dell'atmosfera.
La produzione di O 2
da parte dei Cianobatteri nel volgere
di due miliardi di anni ha modificato la composizione dell'atmosfera che prima
era priva di questo gas. I dati geologici sulla composizione chimica dei
sedimenti indicano che intorno a un miliardo e mezzo di anni fa la
concentrazione di O2 ha
raggiunto in molti luoghi del pianeta una concentrazione pari circa a un
centesimo di quella attuale. A questa concentrazione per le cellule
diventa possibile e conveniente la
respirazione che ha un
rendimento diciotto volte maggiore di quello della fermentazione.
Si presume che le prime cellule eucariotiche
siano comparse appunto circa 1500 milioni di anni fa e che da quel momento le
alghe e poi le piante abbiano contribuito anch'esse a modificare
l'atmosfera. Circa 500 milioni di anni fa nell'atmosfera l'ossigeno ha
raggiunto i valori attuali: un quinto circa in volume, quasi un quarto in
peso. Una massa enorme, calcolata in 5100 miliardi di tonnellate, tuttavia
questa massa non è stabile, si calcola che ogni 5000-6000 anni tutto
l'ossigeno molecolare esistente venga riciclato dagli organismi viventi.
Purché, naturalmente, non intervengano cospicui aumenti del consumo di questo
gas per effetto di combustioni di materiali fossili, nonché riduzione della
sua produzione a sèguito della distruzione dei vegetali fotosintetici.
Flusso di energia e flusso di informazione.
L'energia oltre ad azionare la motilità e la
sintesi di composti organici della cellula, svolge un secondo ruolo nella
fisiologia degli organismi viventi: quella di convogliare l'informazione
relativa alle condizioni che vigono all'esterno dell'organismo, agendo sui
recettori di senso. Questa azione è alla base del capitolo della fisiologia
che concerne la cosiddetta 'vita di relazione'.
I recettori di un organismo hanno compiti
complessi, qui verrà presa in considerazione soltanto la loro funzione di
'trasduttori di energia' grazie alla quale trasformano energia luminosa,
elettrica, chimica, meccanica, termica in segnali atti a dirigere il
comportamento dell'organismo. In altre parole, verrà considerato solo il
loro ruolo di sensori .
Una teoria generale sul ruolo dei sensori in
biologia manca, si può tuttavia dire che gran parte dell'informazione sulle
condizioni esterne proviene dall'azione che una certa forma di energia
esercita sui sensori specifici, cioè sulle strutture atte a trasformarla.
La luce decompone un fotopigmento, l'energia
meccanica deforma una struttura della cellula, una variazione di temperatura
modifica una porzione della membrana plasmatica: in tutti questi casi varia il
potenziale di membrana e viene generato un segnale elettrico che
l'organismo può elaborare ed utilizzare per reagire.
Talvolta i sensori segnalano la presenza di
una sostanza chimica inerte, il benzene ad esempio. Ciò sembra contraddire
l'affermazione appena fatta, ma non è difficile persuadersi che anche in
questo caso si verifica una variazione di potenziale che genera un segnale.
Le cose si svolgono pressappoco così: in
ogni cellula esistono molecole enzimatiche che pompano certi ioni verso
l'esterno e altri verso l'interno e ciò produce una differenza di potenziale
transmembrana di alcuni millivolt (v. § 4.6). Le molecole inerti, ma di forma
e dimensioni adatte, possono intasare i varchi attraverso i quali transitano
questi ioni sicché, anche in questo caso, compare una variazione di potenziale
che può essere trasmessa come segnale interpretabile come presenza della
molecola in questione.
Su questo meccanismo, del resto poco
indagato, non vi è accordo completo. Resta comunque vero che sensori che
rivelano la presenza di sostanze chimiche sono comparsi già presso gli
organismi più antichi e più semplici e che i segnali da essi prodotti dirigono
il movimento della cellula verso la maggiore concentrazione della sostanza,
tassìe positive, o verso la minore concentrazione, tassìe negative.