Derivazione delle monocotiledoni dalle dicotiledoni
Sui rapporti filogenetici tra le due classi del phylum il discorso può procedere lineare e convincente. Le due classi differiscono infatti, non per alcuni pochi caratteri chiave, bensì per una somma di caratteri: rari e sfumati in una di esse, frequenti e pronunciati nell'altra.
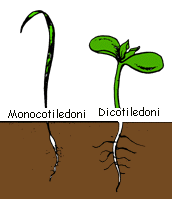
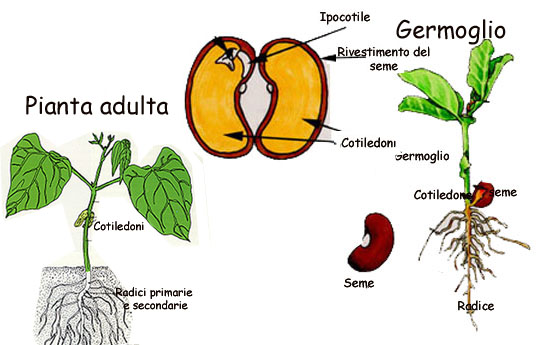
La differenza - che ha dato origine al nome delle classi- riguarda i cotiledoni. Questi organi embrionali (fig.) nel primo caso sono duplici e a forma di foglia, spesso carichi di riserve e capaci di fotosintesi; essi provvedono anche all'assorbimento delle sostanze accumulate nell'endosperma. Nel secondo caso sono singoli e provvedono solo all'assorbimento dell'endosperma. Tuttavia tra le dicotiledoni non mancano, soprattutto presso le ranuncolacee,
semi nei quali i due minuti cotiledoni sono concresciuti.
Altre differenze riguardano talune specializzazioni anatomiche delle monocotiledoni: la radice primaria sparisce e viene sostituita da una corona di radici avventizie. Nel fusto manca la fascia meristematica (cambio) interposta tra xilema e floema (fig.)
che assicura l'accrescimento in larghezza del fusto stesso; questo è invece
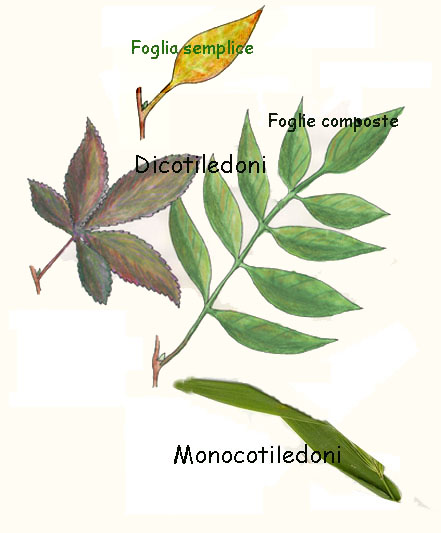
percorso da innumerevoli fasci cribro-vascolari (fig.). Anche le foglie delle monocotiledoni sono particolari poichè
hanno spesso venature parallele e la loro larga base abbraccia il fusto (fig.).
Il fiore delle Monocotiledoni è tripartito ed è costituito da due verticilli di tre tepali ciascuno; ciò si osserva senza difficoltà nel fiore regolare del tulipano e della tradescanzia, e con molta difficoltà nel fiore asimmetrico delle orchidee (fig.).
La struttura tripartita del fiore, i processi sessuali, la fecondazione e anche la morfologia del polline che caratterizzano le Monocotiledoni, insieme a considerazioni comparative embriologiche e anatomiche, indicano una sicura parentela dell'intera classe con le forme erbacee delle Policarpiche e in particolare con le
Ranuncolacee e le Ninfeacee. Questa parentela è tuttavia remota, poiché rimonta almeno al Cretaceo medio-superiore, epoca per la quale
si hanno reperti di palme fossili e di pollini riferibili con sicurezza alle Monocotiledoni.
Le caratteristiche anatomiche qui poste in evidenza sono strettamente correlate e cooperano all'ottimizzazione del flusso dei fluidi nei fasci vascolari che collegano le foglie con le radici e viceversa. Ciò è in rapporto con la rapida crescita e l'abito erbaceo assai comuni presso queste piante che per tal motivo manifestano una pronunciata 'vocazione acquatica': due dei dieci ordini comprendono solo specie acquatiche o palustri, presso altri tre ordini le specie
palustri sono molto frequenti.
Sempre per lo stesso motivo nessuna monocotiledone si difende dal freddo o dalla siccità mediante perdita di foglie, evento tanto comune presso le Dicotiledoni, casomai muore l'intero apparato vegetativo e sopravvivono i
rizomi o i
bulbi.
Meno comune l'adattamento ai climi aridi che viene ottenuto grazie alle foglie succulente coperte di cera, cosa evidente nell'aloe sudafricana e nell'agave del subdeserto messicano ( e divenuta comune in Italia meridionale e nelle isole).
Molti generi di Monocotiledoni grazie alla loro attrezzatura fisiologica hanno potuto colonizzare ambienti proibiti a tutte o quasi le altre piante: le spiagge, i fondali marini, le oasi del deserto, le paludi. A causa di ciò molte specie tendono a formare vastissime comunità monospecifiche: si pensi alle praterie sottomarine di zostere e posidonie, ai palmeti di noci di cocco delle isole oceaniche, ai canneti (fragmiteti) dei delta
fluviali. Tali comunità offrono cibo e riparo ad una fauna specializzata, peraltro soffrono dei rischi delle monocolture praticate dall'uomo: una malattia epidemica che provochi estese
morie di questi organismi fotosintetici ('produttori primari') può annientare temporaneamente ogni forma di vita in aree molto vaste.