|
Le lamprede (fig) sono diffuse nelle acque interne e
nei mari dei paesi a clima temperato, sono quindi comuni nei paesi del
Mediterraneo. Gli antichi Romani ne apprezzavano le carni, ma oggi le lamprede
valgono poco anzi, si lamentano i danni che recano ai pesci dei Grandi Laghi del
Nordamerica che esse dissanguano diminuendone il valore sui mercati (fig.).
Biologia e comportamento
Le lamprede nascono, crescono e si riproducono nelle acque dolci,
ma molte specie completano la crescita in mare dove divengono parassite dei
pesci ai quali si attaccano con la ventosa che circonda la bocca, e quindi
succhiano il loro sangue. Le lamprede risalgono poi i fiumi sino a raggiungere i
ruscelli di montagna senza più nutrirsi e nei luoghi idonei si accoppiano con
fecondazione interna; dopo la deposizione delle uova fecondate esse muoiono. E’
questo un ciclo vitale unico presso i vertebrati che di regola si riproducono
più volte nel corso della vita adulta.
Dalle uova, che hanno il diametro di circa un
millimetro, schiudono larve che presto vengono a somigliare per modo di vivere e
per morfologia, all’ anfiosso. Stanno sepolte
nel fango ma affiora la loro bocca che è fornita di brevi appendici. Pompano
l’acqua per mezzo dei sacchi branchiali provvisti di robuste pareti muscolari (fig).
Il flusso d’acqua trasporta fine materiale organico e minuscoli organismi che
vengono invischiati nel muco prodotto dall’ endostilo, trascinati nell’intestino
e quindi digeriti. Vivono 5-7 anni in questo modo e, quando hanno raggiunto le
dimensioni definitive, metamorfosano in adulti muniti di efficienti organi di
senso e di gonadi. I giovani adulti delle specie anadrome si dirigono
verso il mare, lì si nutrono e accrescono e poi risalgono verso i luoghi di
riproduzione, mentre gli adulti delle specie non migranti si riproducono sul
posto.
In definitiva, alle larve di lamprede compete il
nutrirsi e l’accrescersi, mentre alla più breve fase adulta compete, oltre al
riprodursi, la panmissia e la dispersione in luoghi lontani. Simile
strategia, che ha risolto problemi gravi per tutti gli animali dulciacquicoli,
ha consentito a questi vertebrati primitivi di sopravvivere per centinaia di
milioni di anni e persino di resistere all’inquinamento ambientale causato dalla
tecnologia umana poiché, se il corso d’acqua da cui provenivano è diventato
invivibile ne scelgono uno migliore. Identica strategia è evoluta presso molti
insetti volatori che hanno larve acquatiche: gli
adulti degli Efemerotteri, dei
Tricotteri e di molti Ditteri,
sfarfallano e, senza nutrirsi, provvedono a disperdersi e riprodursi.
Anatomia funzionale delle
lamprede
La larva delle lamprede somiglia all’ anfiosso
(fig),
tuttavia presenta differenze cospicue per quanto riguarda la regione del capo e
la regione branchiale. Entrambe le regioni sono sostenute da uno scheletro
cartilagineo: sulla conca cartilaginea anteriore poggia l’encefalo, ancora non
del tutto sviluppato, connesso con gli organi di senso olfattivo, stato-acustico
e con gli occhi anch’essi rudimentali (fig. ).
La corda dorsale non si spinge fino all’apice anteriore della
larva, ma si arresta a metà circa dell’encefalo.
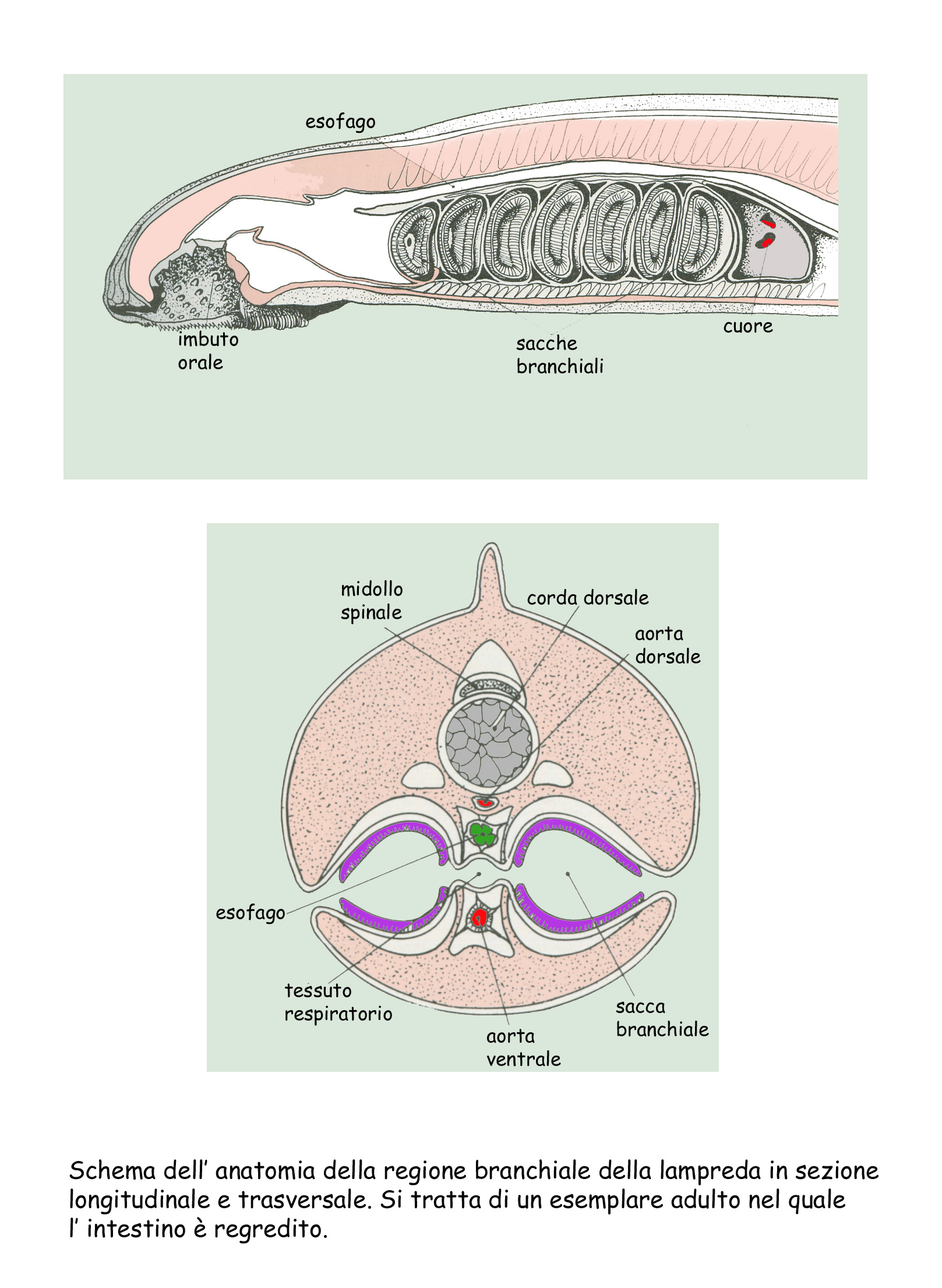
Le branchie sono racchiuse entro sacchi dalle
pareti muscolose. Questi sacchi si contraggono spingendo l’acqua che contengono
verso l’esterno, poi si rilasciano e, grazie all’ elasticità delle cartilagini,
aspirano l’acqua attraverso la bocca. Questo apparato è molto più efficiente nel
generare un flusso d’acqua, contenente ossigeno e nutrimento, di quanto non sia
l’apparato branchiale dell’anfiosso coperto da ciglia vibratili. Pertanto si
osserva che le branchie della larva della lampreda sono molto meno numerose di
quelle dell’anfiosso e che la faringe è più breve. Di fronte ai sacchi
branchiali vi è l’endostilo
impari.
L’apparato circolatorio è chiuso ed ha come
motore il cuore. Questo, essendo collegato a una capsula pericardica, solida ma
elastica, si può dilatare dopo la sistole e aspira così il sangue venoso.
Nell’adulto, che conduce vita molto più attiva,
si completa lo sviluppo degli organi di senso: gli occhi laterali raggiungono la
struttura tipica per i vertebrati, mentre il terzo occhio situato dorsalmente
presso l’epifisi non cambia di struttura e rimane molto piccolo e semplice, l’apparato branchiale migra in un diverticolo e l’endostilo si trasforma in una
ghiandola a secrezione interna, la tiroide (fig.).
Avanti al terzo occhio si apre una narice impari
munita di un condotto che raggiunge la sacca olfattoria; dal condotto si stacca
un diverticolo a fondo cieco che si pone in contatto con l’ipofisi, ghiandola
situata sotto il diencefalo, che produce ormoni che governano la riproduzione e
il metabolismo dell’animale.
|
Classi e ordini degli Agnati
Petromizontiformi
Missiniformi
Regione branchiale della lampreda in sezione longitudinale e
trasversale
Disposizione dei sacchi branchiali in due
Missine
|
I Missinoidi oggi viventi differiscono dai Petromizonti per avere la narice impari con apertura anteriore e sbocco entro la
faringe in prossimità dell’ ipofisi. Le pinne dorsale e anale sono rudimentali e
più spesso assenti, quelle pari mancano sempre (fig. ). Lo scheletro
cartilagineo è poco sviluppato e, in particolare, il cervello è sostenuto da un
semplice foglio membranoso, anche la capsula pericardica manca e il ritorno del
sangue venoso al cuore è assicurato da cuori ausiliari situati verso la coda.
Nelle forme fossili lo scheletro cutaneo è formato da cristalli di un tipo di
fosfato di calcio denominato idrossiapatite.
Le branchie sono racchiuse entro sacchi globulari che da una
parte si aprono nella faringe e dall’ altra sfociano all’ esterno con canali più o meno lunghi che tendono a confluire su ciascun lato
(fig. ). L’apparato branchiale funziona solo per gli scambi gassosi e l’
alimentazione per filtrazione è sostituita dalla cattura di prede più o meno
inerti che vivono nella sabbia e nella fanghiglia dei fondi marini, oppure dalla
consumazione di cadaveri di più grossi animali. A questi fini le missine
adoperano la ‘lingua’ costituita da un lungo muscolo fusiforme che porta in cima
una serie di dentelli.
Gli occhi e l’ apparato statoacustico sono
rudimentali, anche l’apparato della linea laterale è rudimentale o, più spesso,
assente.
La differenza più cospicua tra Petromizonti e
Missinoidi non riguarda l’ anatomia, ma la fisiologia. Riguarda il fatto che i
fluidi interni sono isotonici con l’ acqua marina, diversamente da quanto accade
per tutti gli altri vertebrati i cui fluidi interni hanno pressione osmotica
pressappoco intermedia tra quelle del mare e quella d’ acqua dolce e sono, di
regola, sotto attivo controllo omeostatico che fa capo all’ipofisi. I Missinoidi
per tale fatto sono, e quasi sicuramente sono sempre stati, strettamente marini.
Le uova delle missine sono fusiformi e
piuttosto grandi, misurano circa 10 mm di lunghezza. Lo stadio larvale manca in
tutte le specie dei Missinoidi viventi e si hanno prove che mancasse anche in
quelle fossili.
Adattamenti funzionali degli Agnati petromizonti
fossili.
Animali
simili a Myllokunmingia sono classificati in vari generi dell’ordine
degli Anaspidi, nome che significa ‘privi di scudo’. Gli Anaspidi si
succedono per molte decine di milioni di anni fino a tutto il Devoniano e dànno
origine a ceppi poco diversi. In un ceppo il corpo era coperto da squame rade e
leggere; in un altro da squame ben embricate di fosfato di calcio; queste squame
oltre alla funzione protettiva funzionavano da zavorra e permettevano all’
adulto di nutrirsi presso il fondo col pulviscolo di detriti che si sollevava
davanti a lui quando nuotava rasente il fondo. Le due pinne longitudinali erano sostenute da
solide spine triangolari.
Gli adulti di un altro gruppo, quello dei
Cefalaspidi (fig), che più degli altri si era inoltrato nelle acque interne, era
munito di una pesante corazza che racchiudeva il capo e la parte anteriore del
corpo. Le forme meno evolute presentano nell’ adulto le aperture branchiali sui
fianchi (Tremataspis), ma nelle forme successive queste aperture
si presentano spostate ventralmente (fig.). In entrambi i casi il margine
dello scudo, sede delle aperture branchiali, sosteneva un pavimento flessibile
formato da piccole placche separate. Quando l’ animale veniva disturbato o
minacciato da un predatore - gli scorpioni d’acqua (Euripteridi) avevano invaso
le acque dolci - allora i muscoli, contraendosi, sollevavano il pavimento verso
la volta della corazza cefalica e l’ acqua contenuta nelle branchie veniva
espulsa con forza verso il basso. Per reazione l’ animale riceveva una spinta
verso l’ alto e quindi poteva mettersi a nuotare entro una nuvola di fango,
orientandosi nel buio grazie al campo elettrico generato dagli organi elettrici
situati sul capo.
Questa ricostruzione del comportamento dei Cefalaspidi, che può apparire un po’ fantasiosa e gratuita, si vale degli studi
di Stensiö, paleontologo svedese, il quale, grazie a una raffinata tecnica, è
riuscito a ricostruire i fini dettagli anatomici del loro cervello e dei loro
nervi cranici, compreso il nervo vago, che innerva appunto gli organi elettrici.
Va anche aggiunto che questi organi funzionano al meglio in acqua dolce a bassa
conduttività piuttosto che in acqua marina ad alta conduttività, in questo caso
il campo elettrico non è stabile e si dilegua prontamente(fig).
Origine ed evoluzione dei Ciclostomi
Gli Anaspidi (fig) e i Cefalaspidi sono scomparsi nel corso del
Devoniano, periodo in cui nelle
acque marine e in quelle dolci si affollavano i loro discendenti, meglio
attrezzati con mascelle munite di denti e robuste pinne pari. Si è però salvata
una terza sottoclasse dei Petromizonti, denominata Ciclostomi, i cui primi
fossili sono stati rinvenuti nei giacimenti del Carbonifero. Di questo gruppo
sopravvivono tuttora una trentina di specie.
I Ciclostomi differiscono dagli Anaspidi, dai quali si presume
che siano derivati, per aver perso del tutto il rivestimento di squame e le due
lunghe pinne ventro-laterali; queste perdite sono da porre in rapporto con la
struttura anguilliforme che essi hanno acquistato. Infatti anche le anguille
attuali, che sono veri pesci, sono prive di squame e pinne pari.
I Ciclostomi hanno inoltre acquistato una ventosa circolare che
circonda la bocca (fig). Questa ventosa, come la ventosa
delle sanguisughe, ha varie funzioni: quella di attaccarsi al substrato solido
quando l’ animale si trova in acque turbolente è probabilmente primitiva; viene
poi quella di attaccarsi ai pesci abili nuotatori per farsi trasportare senza
spesa; viene infine quella di succhiare il sangue del trasportatore, utilizzando
come stiletto la lingua armata di dentini.
La lampreda, comunque, si fa trasportare solo verso il mare, il
ritorno lo compie con le proprie forze.
Le modifiche fisiologiche che permettono alle lamprede di
lasciare l’ambiente dulciacquicolo per affrontare quello marino sono controllate
dalla prolattina, ormone secreto dall’ipofisi, presente in tutti i vertebrati,
ma con funzioni diverse sempre connesse con la riproduzione.
Per quanto riguarda l’ evoluzione molecolare è interessante
notare che nelle lamprede le molecole dell’
emoglobina sono formate da
semplici dimeri anziché da tetrametri; altrettanto vale per le molecole delle
immunoglobuline. Si tratta di condizioni primitive che si sono conservate per
centinaia di milioni di anni.
Nota: La
sistematica degli Agnati è ancora fluida, qui si propone una classificazione
provvisoria che contempera
le concezioni di paleontologi e di zoologi.
| | |

{kind=link}