Al di sopra della bocca si articola un labbro superiore, mentre mento e submento formano il labbro inferiore (fig.).
Il nome di queste parti buccali, sia chiaro, indica solo che esiste una vaga analogia con le strutture di egual
nome dei vertebrati; del resto esse sono presenti e riconoscibili soltanto in
quegli insetti che si nutrono rodendo e masticando il loro cibo: blatte,
cavallette, carabi, mantidi,vespe, formiche e altri ancora. In taluni ordini di
insetti tutte queste strutture sono modificate in rapporto al modo di assumere
il cibo: l'apparato buccale diviene succhiatore nelle farfalle, giunte allo
stato di immagine, pungente negli afidi e cicale, pungente e succhiatore in
zanzare (fig.), pidocchi, cimici, pulci,
lambente nelle comuni mosche. A questo proposito è da notare che presso gli insetti in cui
compaiono larve, queste hanno spesso dieta, e quindi apparato buccale, del tutto diverso da quello dell'adulto o immagine; i casi più vistosi riguardano le pulci, le farfalle e le zanzare (femmine), ma si potrebbero fare moltissimi altri esempi. Da notare infine che gli adulti degli
Efemerotteri, di vari Plecotteri, Lepidotteri e Ditteri, non si alimentano, sicché hanno apparato buccale regredito.
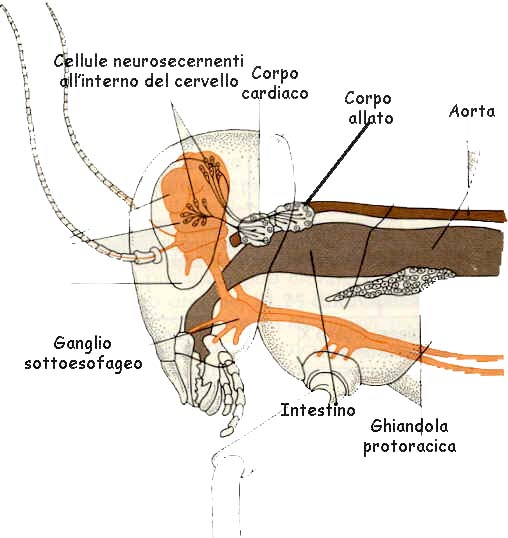
Nella capsula cefalica sono racchiusi i gangli sopraesofagei (cerebro)
nonché tre paia di gangli sottoesofagei che si continuano con la catena nervosa ventrale. Ai gangli sopraesofagei sono annessi due centri neurosecretori che producono un
ormone regolatore dell'intero ciclo vitale e un ormone (ormone giovanile) che controlla la crescita
(fig.). Un terzo ormone steroideo secreto da una
ghiandola protoracica (ecdisone) controlla la muta.
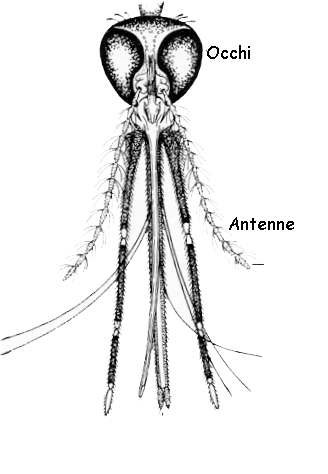
Al sistema nervoso fanno capo i recettori visivi degli occhi composti e degli ocelli, i recettori olfattivi dislocati sulle antenne, i meccanocettori dell'organo di Johnston situati alla base delle antenne,
nonché una miriade di sensilli dislocati
in prossimità della bocca e sulle estremità delle zampe.
(fig)
Il torace degli insetti comprende tre segmenti denominati protorace, mesotorace, metatorace.
Ciascun segmento toracico porta un paio di zampe formate tipicamente da 5 segmenti:
Quest'ultimo è di regola suddiviso in più articoli e termina con due unghie
(presso gli entognati le unghie mancano o ve n'è una) (fig.).
Negli insetti pterigoti il mesotorace e il metatorace portano un paio di ali incernierate tra i tergi e le 'pleure' (fig.). I muscoli agiscono in parte sull'ala, in parte sul tergite con il quale l'ala è incernierata. L'aerodinamica
delle ali è resa complessa dalla varietà dei movimenti che possono venir loro impressi
nonché dallo svergolamento del piano alare e dal diverso comportamento del paio anteriore rispetto al posteriore; tuttavia nel caso che le due ali di ciascun lato siano agganciate esse si comportano come un tutt'uno. In linea di massima l'ala compie un movimento avvolgente col quale sospinge l'aria verso il basso e l'indietro: lo sforzo meccanico si concentra sul bordo anteriore che è
rinforzato da particolari nervature e placche (stigmi), il bordo posteriore è invece libero. Le ali sono duplicature dell'esoscheletro rafforzate da nervature longitudinali e trasversali. Le nervature longitudinali tendono a fondersi o, meno spesso, a ramificarsi; se ne distinguono sette (singole o multiple).
In base alle venature si distinguono quattro tipi di ali, ciascuno dei quali caratterizza una 'serie' di insetti pterigoti (Tab.)
Nel torace sono ospitate tre paia di gangli
nervosi più o meno fusi.
Nei solchi posteriori di ciascun segmento toracico, lateralmente, si aprono le prime due paia di
spiracoli dai quali si diramano altrettante
trachee.
L'addome negli Euentòmati è formato da undici segmenti; va notato però che gli ultimi tendono a sparire, sicché di solito se ne contano meno. Quando l'addome è lungo e le zampe sono brevi, come avviene negli
insetti che vivono in crepacci e sotto i sassi, l'insetto trascina la sua parte posteriore. Nel caso dei Tisanuri l'addome scivola su stili che riducono l'attrito, ma nel caso delle Forficule, dei Coleotteri Stafilinidi e di certe larve esso può essere sollevato e inarcato verso il dorso quando l'animale si trova allo scoperto, ciò rappresenta anche un atteggiamento di minaccia.
Negli insetti volatori l'inserzione delle ali può trovarsi situata innanzi al centro di gravità, in tal caso l'addome pende in giù per non turbare l'assetto di volo.
Le larve degli insetti appartenenti all'ordine degli
Efemerotteri e alla serie degli
Oligoneotteri sono spesso dotate di appendici addominali omologhe alle zampe, di solito non sono adibite alla locomozione
(fig). In questo e in altri ordini
l'ultimo segmento porta i cerci e/o un filo terminale (fig)
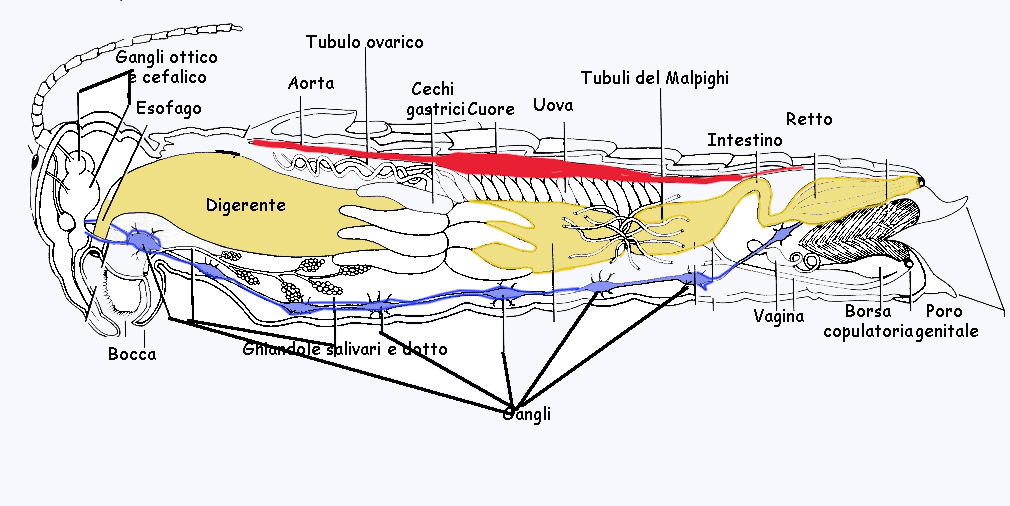
Nell'addome sono contenuti: il vaso dorsale contrattile, la maggior parte dell'apparato tracheale, la maggior parte del tubo digerente, le gonadi, gli organi copulatori, parte della catena nervosa.
Il vaso dorsale ha la stessa struttura e lo stesso funzionamento che ha presso gli
Onicofori e per altri Artropodi: è formato da una serie di concamerazioni ciascuna munita di due aperture, gli ostioli. La dilatazione di ciascuna
concamerazione prodotta dai muscoli aliformi (estrinseci, agganciati all’esoscheletro) aspira l'emolinfa dagli spazi circostanti; la successiva contrazione della muscolatura intrinseca delle pareti del vaso la spinge verso l'avanti entro l'aorta che percorre il torace e giunge fino al capo. A seconda della specie, dall'aorta si diramano alcuni vasi minori che si spingono entro le zampe e le ali, ma queste diramazioni possono mancare, soprattutto negli animali di piccola
taglia. L'emolinfa degli insetti è priva di pigmenti per il trasporto di ossigeno che risultano superflui poichè le trachee che convogliano l'aria si suddividono in ramificazioni finissime portando l'ossigeno ovunque, persino all'interno delle cellule che più ne richiedono.
Il sistema tracheale è collegato sia longitudinalmente, sia trasversalmente, e in molti casi si dilata in sacchi aerei che migliorano il rapporto volume/peso. L'aria ricca di O2 viene inspirata, e quella ricca di CO2 espulsa, attraverso gli spiracoli (o stigmi) di cui ve ne sono, tipicamente, otto paia nell'addome e due paia nel torace; questo numero tende a ridursi, fino a due nel caso delle cocciniglie (famiglia dei Coccidi, ordine degli Omotteri). La ventilazione è assicurata per lo più dalle contrazioni dell'addome che si allunga e contrae come un organetto.
Il tubo digerente degli Insetti si divide in tre porzioni. Quella anteriore (stomodeo) e quella posteriore (proctodeo) sono tappezzate di cuticola chitinosa. Detta cuticola, pur garantendo una certa permeabilità, offre sia una protezione meccanica, sia una barriera contro l'ingresso di microorganismi indesiderati nei tessuti. L'epitelio dell'intestino medio è invece nudo, ma dall'anello di congiunzione tra intestino medio e anteriore si stacca e si
prolunga verso l'indietro un sottile manicotto membranoso, la membrana peritrofica, che ha il medesimo compito protettivo del rivestimento cuticolare. La
digestione negli insetti si svolge tutta nell'intestino, mancano infatti le 'ghiandole' deputate alla digestione intracellulare e mancano anche ghiandole ausiliari per la secrezione di enzimi ad eccezione delle ghiandole salivari che si aprono nella bocca.
La lunghezza dell'intestino è maggiore negli insetti nella cui dieta prevalgono i
polisaccaridi. Va specificato, tuttavia, che la digestione presso gli insetti è coadiuvata da microorganismi simbionti vari (batteri, miceti, protozoi) i
quali, oltre agli enzimi necessari, forniscono anche composti del tipo delle vitamine che l'insetto non è in grado di sintetizzare; questo apporto è indispensabile per gli insetti a dieta povera e uniforme, in particolare per quelli che si nutrono di cellulosa e
lignina, di cheratina, e anche per quelli che succhiano sangue. Di regola è la madre che, prima di deporlo, dà in dotazione all’uovo i microorganismi
simbionti.
Al confine tra intestino medio e intestino posteriore si diramano i tubuli
malpighiani (fig.), che hanno funzione escretrice e riversano le sostanze di rifiuto nel tratto posteriore dell'intestino.
Le gonadi (fig.) sono pari. I testicoli sono formati da sottili tubuli ramificati, ove vengono prodotti gli spermi. Lungo lo spermidotto si trovano una o due vescicole seminali ove si accumulano gli spermi che verranno emessi durante
l'accoppiamento.
Gli ovari sono costituiti da fasci di ovarioli conformati in modo diverso a seconda del modo in cui le uova vengono rifornite di tuorlo; le cellule follicolari che circondano le uova forniranno a queste un involucro speciale
chiamato corion. Il corion presenta il micropilo, porticina di ingresso per lo
spermatozoo.
Il grado di protezione contro la disidratazione offerta dal corion varia molto presso i diversi ordini, ciò condiziona la scelta del luogo in cui la femmina deporrà le sue uova: in acqua o dentro i tessuti vegetali o animali nelle specie con uova dotate di involucri permeabili, entro il terriccio o in nidi ben protetti per quelle specie le cui uova sono mediocremente protette, all'aperto per quelle che depongono uova ben impermeabilizzate. Questo è il caso delle
farfalle, dei fasmidi, dei pidocchi, delle pulci e di non molti altri insetti; la struttura del corion in queste specie è molto complessa poichè dev'essere impedita la fuoriuscita di vapor acqueo e nello stesso tempo consentito l'ingresso dell'ossigeno indispensabile all'embrione in sviluppo.
La fecondazione è sempre interna, ma variano le
modalità della trasmissione degli spermi. Gli sbocchi delle gonadi sono situati
tra 8° e 9° segmento: in alcuni ordini la femmina è provvista di un ovopositore che consente la deposizione
delle uova ad una certa profondità nel terreno o entro i tessuti di piante o di animali.
Negli Insetti Euentòmati la morfogenesi può seguire strade diverse (fig.).
Lo sviluppo è diretto negli Apterigoti il cui giovane schiude dall'uovo con le medesime caratteristiche dell'adulto: durante il susseguirsi delle mute crescono solo le dimensioni dell'individuo e variano i rapporti tra le diverse parti.
Nella maggior parte dei Polineotteri e dei Paraneotteri il giovane nasce invece privo di ali ed equipaggiato per la locomozione terrestre (o per il nuoto) in modo non diverso dall'adulto; acquisterà la capacità di volare quando ha effettuato
l'ultima muta allo scopo soprattutto di trovare un partner e il luogo più idoneo per la deposizione delle uova. Il passaggio dalla condizione giovanile a quella adulta è graduale e gli insetti che così si sviluppano vengono anche chiamati eterometaboli; gli individui giovanili sono indicati col nome di neànidi, l'adulto col nome di immagine.
Nei Paleotteri che conducono vita anfibia e in quei Polineotteri che vivono allo stesso modo, gli stadi preimmaginali sono equipaggiati con strutture di tipo branchiale che spariranno negli adulti volatori. Il passaggio allo stadio adulto
avviene quindi in modo più brusco e gli insetti che così si sviluppano vengono detti emimetaboli. Negli Oligoneotteri lo sviluppo procede in modo diverso: gli stadi preimmaginali, detti anche larvali, non somigliano allo stadio adulto, né per la forma, né per l modo di alimentarsi, né per l'ambiente occupato, e nemmeno per equipaggiamento sensoriale. Le differenze sono tanto forti che per gli insetti oligoneotteri non è possibile attribuire
larva e adulto alla stessa specie, a meno che non si sia assistito allo sfarfallamento'(fig.) Lo stadio che precede lo sfarfallamento viene indicato col nome di pupa, o di crisalide nel caso delle farfalle. La pupa è immobile poiché in essa ha luogo il riassorbimento di quei tessuti che risulteranno superflui all'adulto e l'edificazione delle strutture proprie della immagine. Gli insetti che così si sviluppano vengono detti olometaboli,
termine che indica la totale ristrutturazione dell'organismo.
Il fatto che il medesimo patrimonio ereditario
possa attuare strutture radicalmente diverse a sèguito di un segnale endocrino
piuttosto semplice, rappresenta per la genetica e per l'evoluzionismo un grosso
problema da risolvere.


{kind=link}
{kind=link}